- Current
- Browse
- Collections
-
For contributors
- For Authors
- Instructions to authors
- Article processing charge
- e-submission
- For Reviewers
- Instructions for reviewers
- How to become a reviewer
- Best reviewers
- For Readers
- Readership
- Subscription
- Permission guidelines
- About
- Editorial policy
Articles
- Page Path
- HOME > Diabetes Metab J > Volume 43(5); 2019 > Article
-
ReviewOthers Mitochondrial Toxins and Healthy Lifestyle Meet at the Crossroad of Hormesis
-
Yu-Mi Lee1
 , Duk-Hee Lee1,2
, Duk-Hee Lee1,2 -
Diabetes & Metabolism Journal 2019;43(5):568-577.
DOI: https://doi.org/10.4093/dmj.2019.0143
Published online: October 24, 2019
1Department of Preventive Medicine, School of Medicine, Kyungpook National University, Daegu, Korea.
2BK21 Plus KNU Biomedical Convergence Program, Department of Biomedical Science, Kyungpook National University, Daegu, Korea.
- Corresponding author: Duk-Hee Lee. Department of Preventive Medicine, School of Medicine, Kyungpook National University, 680 Gukchaebosang-ro, Jung-gu, Daegu 41944, Korea. lee_dh@knu.ac.kr
• Received: July 19, 2019 • Accepted: August 11, 2019
Copyright © 2019 Korean Diabetes Association
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
- ABSTRACT
- INTRODUCTION
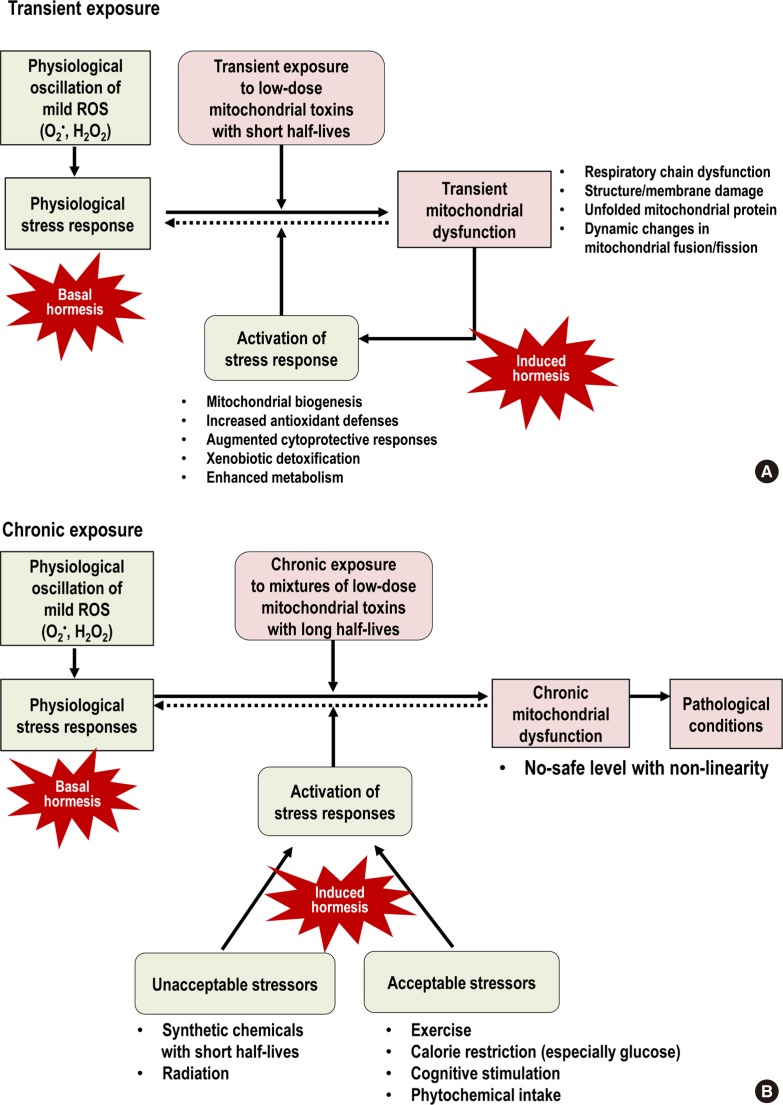
- MITOCHONDRIA STRESS RESPONSE
- PERSISTENT CHEMICAL MIXTURES AS MITOCHONDRIAL TOXINS
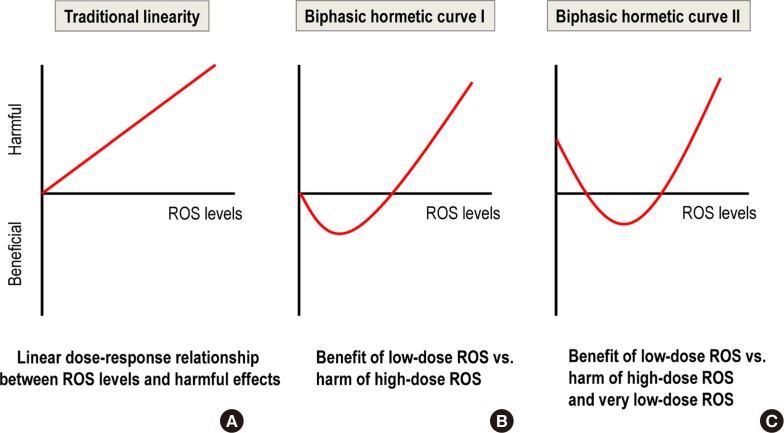
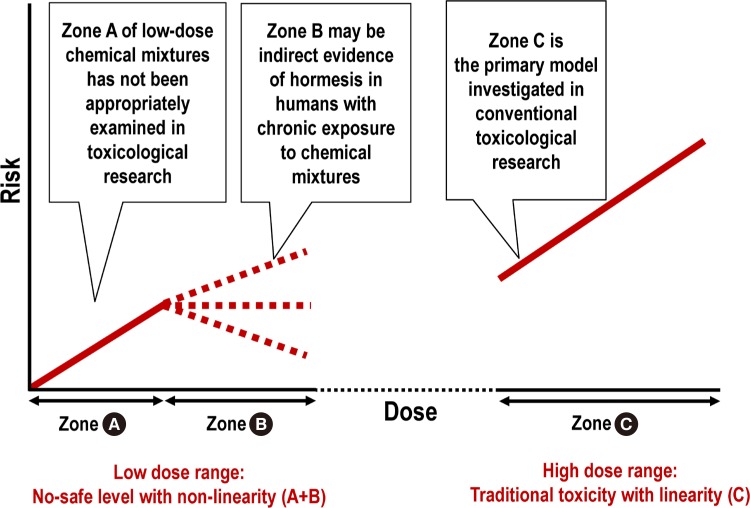
- HORMESIS CAN EXPLAIN THE NON-LINEAR RESPONSE IN THE LOW-DOSE RANGE
- CURRENT APPROACHES CANNOT EFFECTIVELY PROTECT HUMANS FROM THE HARM OF LOW-DOSE ENVIRONMENTAL CHEMICALS
- MITOCHONDRIAL TOXINS AND HEALTHY LIFESTYLES CAN MEET AT THE CROSSROAD OF HORMESIS
- CONCLUSIONS
- ACKNOWLEDGMENTS
- NOTES
- REFERENCES
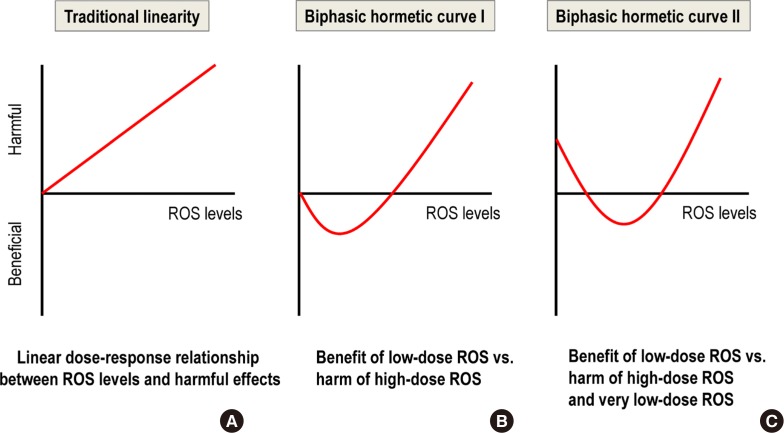
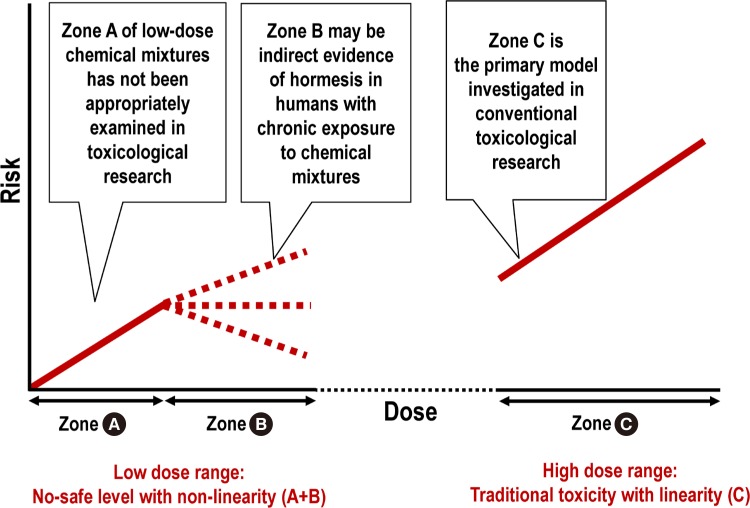
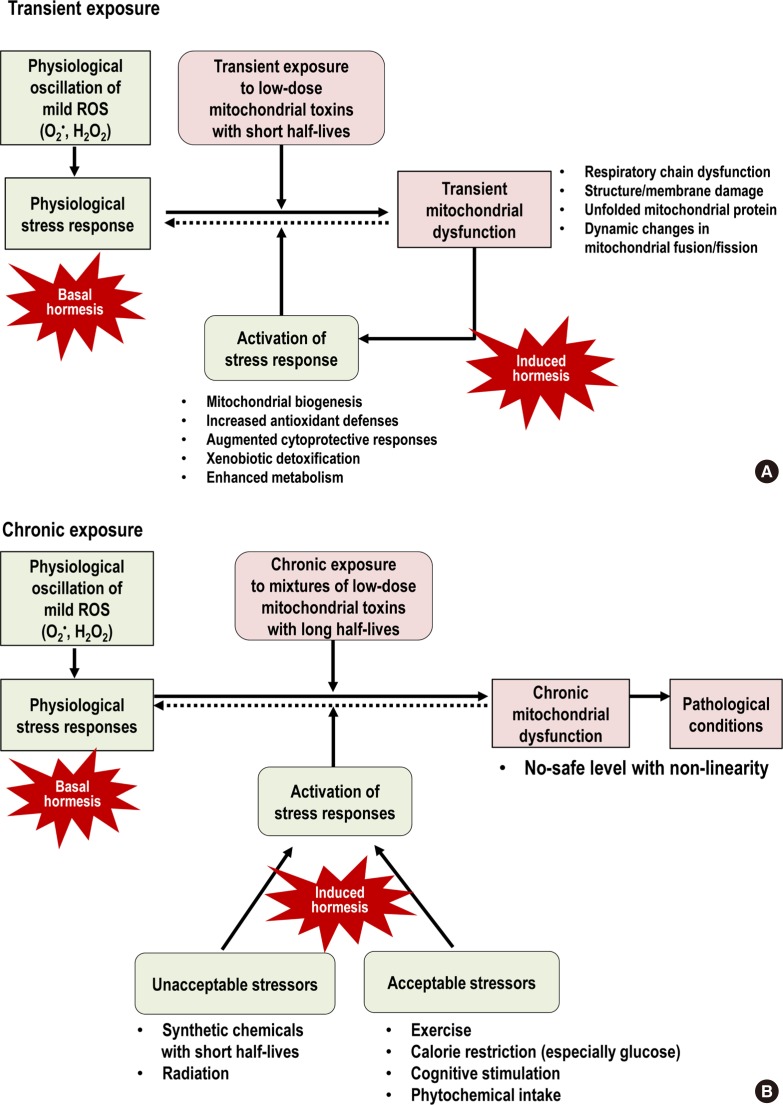
Figure & Data
References
Citations
Citations to this article as recorded by 
- Polyethylene terephthalate (PET) micro- and nanoplastic particles affect the mitochondrial efficiency of human brain vascular pericytes without inducing oxidative stress
Sean M. Gettings, William Timbury, Anna Dmochowska, Riddhi Sharma, Rebecca McGonigle, Lewis E. MacKenzie, Guillaume Miquelard-Garnier, Nora Bourbia
NanoImpact.2024; 34: 100508. CrossRef - Rules of Heliogeomagnetics Diversely Coordinating Biological Rhythms and Promoting Human Health
Kuniaki Otsuka, Germaine Cornelissen, Andi Weydahl, Denis Gubin, Larry A. Beaty, Masatoshi Murase
Applied Sciences.2023; 13(2): 951. CrossRef - Can lipophilic pollutants in adipose tissue explain weight change‐related risk in type 2 diabetes mellitus?
Duk‐Hee Lee, In‐Kyu Lee
Journal of Diabetes Investigation.2023; 14(4): 528. CrossRef - Hormetic Effects of Cerium, Lanthanum and Their Combination at Sub-micromolar Concentrations in Sea Urchin Sperm
Giovanni Pagano, Antonios Apostolos Brouziotis, Daniel Lyons, Ivana Čarapar, Rahime Oral, Serkan Tez, Philippe J. Thomas, Franca Tommasi, Giovanni Libralato, Marco Guida, Marco Trifuoggi
Bulletin of Environmental Contamination and Toxicology.2023;[Epub] CrossRef - Mitochondria: It is all about energy
Amaloha Casanova, Anne Wevers, Santiago Navarro-Ledesma, Leo Pruimboom
Frontiers in Physiology.2023;[Epub] CrossRef - Type 2 Diabetes Induced by Changes in Proteomic Profiling of Zebrafish Chronically Exposed to a Mixture of Organochlorine Pesticides at Low Concentrations
Yan Gao, Hyojin Lee, Sangkyu Lee, Ki-Tae Kim
International Journal of Environmental Research and Public Health.2022; 19(9): 4991. CrossRef - Effect of Low-Dose Persistent Organic Pollutants on Mitochondrial Function: Human and in Vitro Evidence
Se-A Kim, Hoyul Lee, Sung-Mi Park, Mi-Jin Kim, Yu-Mi Lee, Young-Ran Yoon, Hyun-Kyung Lee, Hyo-Bang Moon, In-Kyu Lee, Duk-Hee Lee
Diabetes & Metabolism Journal.2022; 46(4): 592. CrossRef - Can Environmental Pollutants Be a Factor Linking Obesity and COVID-19?
Duk-Hee Lee
Journal of Korean Medical Science.2021;[Epub] CrossRef - Intensive weight loss and cognition: The dynamics of persistent organic pollutants in adipose tissue can explain the unexpected results from the Action for Health in Diabetes (Look AHEAD) study
Yu‐Mi Lee, Sun‐Hee Park, Duk‐Hee Lee
Alzheimer's & Dementia.2020; 16(4): 696. CrossRef - Lipophilic Environmental Chemical Mixtures Released During Weight‐Loss: The Need to Consider Dynamics
Duk‐Hee Lee, David R Jacobs, Lars Lind, P. Monica Lind
BioEssays.2020;[Epub] CrossRef - Environmental toxicology and ecotoxicology: How clean is clean? Rethinking dose-response analysis
Evgenios Agathokleous, Edward J. Calabrese
Science of The Total Environment.2020; 746: 138769. CrossRef
 PubReader
PubReader Cite
Cite