- Current
- Browse
- Collections
-
For contributors
- For Authors
- Instructions to authors
- Article processing charge
- e-submission
- For Reviewers
- Instructions for reviewers
- How to become a reviewer
- Best reviewers
- For Readers
- Readership
- Subscription
- Permission guidelines
- About
- Editorial policy
Articles
- Page Path
- HOME > Diabetes Metab J > Volume 48(1); 2024 > Article
-
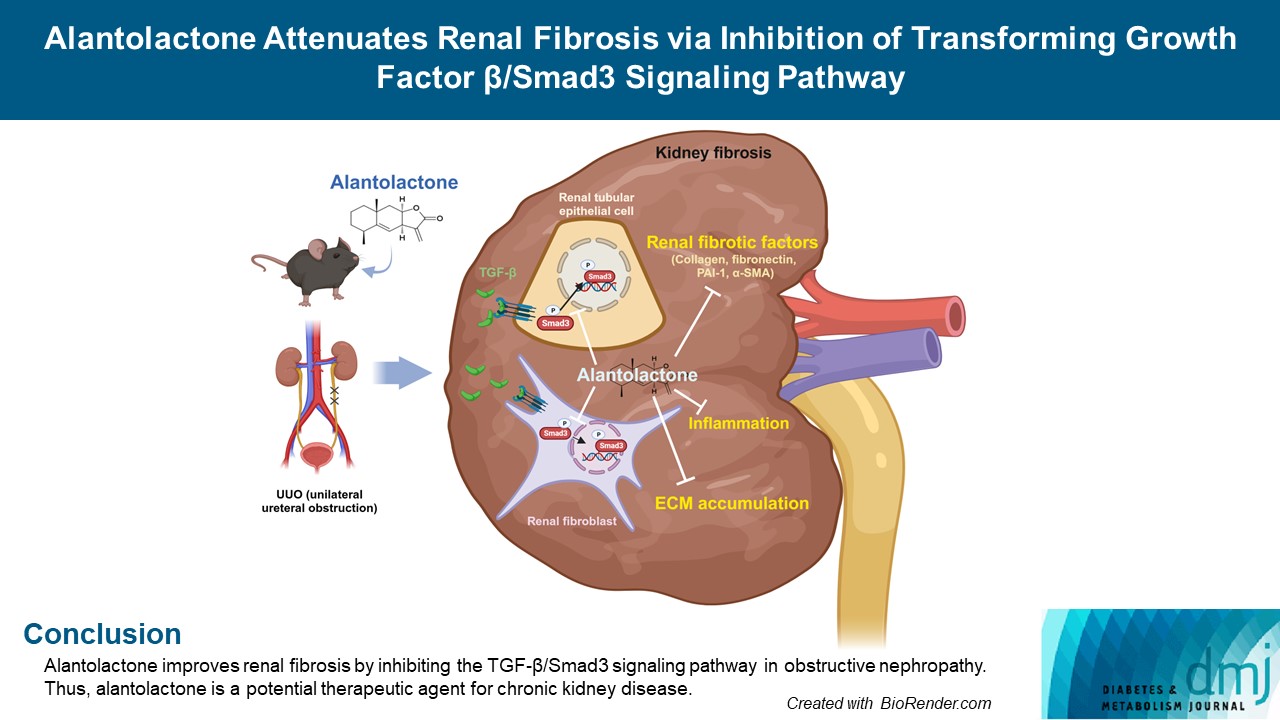
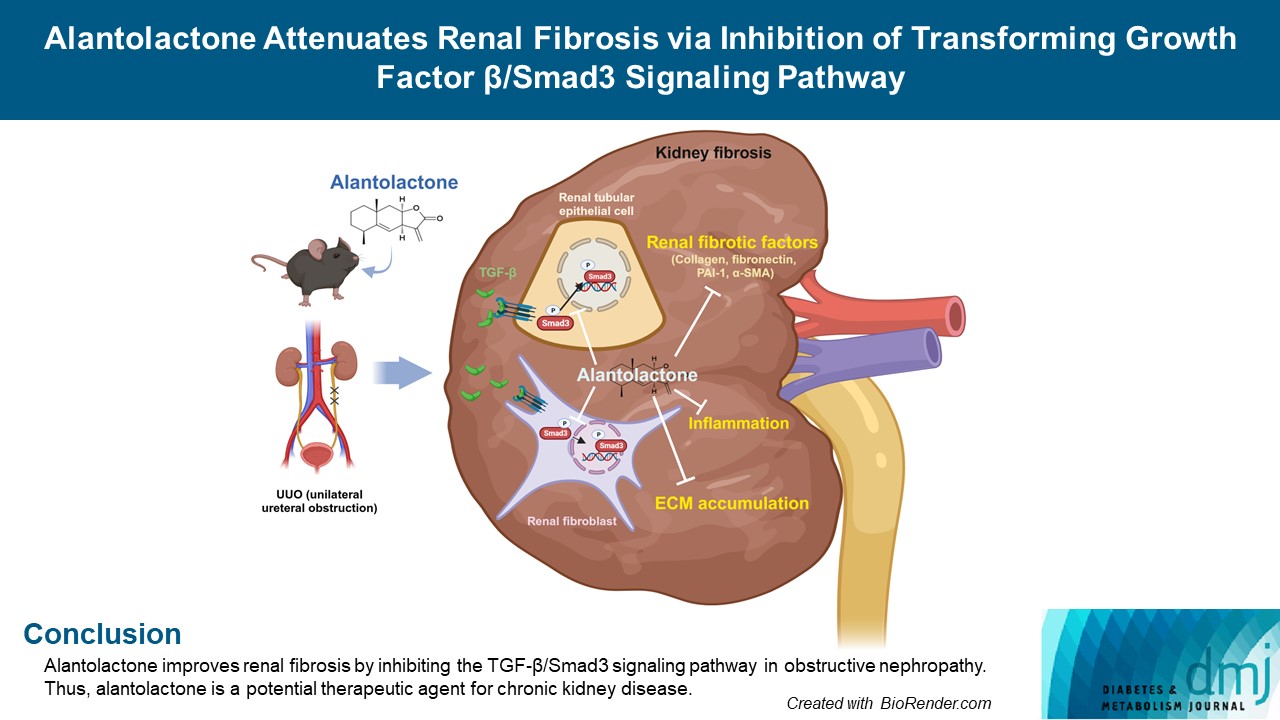
Original ArticleBasic Research Alantolactone Attenuates Renal Fibrosis via Inhibition of Transforming Growth Factor β/Smad3 Signaling Pathway
-
Kyeong-Min Lee1,*
 , Yeo Jin Hwang1*, Gwon-Soo Jung2
, Yeo Jin Hwang1*, Gwon-Soo Jung2 -
Diabetes & Metabolism Journal 2024;48(1):72-82.
DOI: https://doi.org/10.4093/dmj.2022.0231
Published online: January 3, 2024
- 1,342 Views
- 146 Download
1Division of Biotechnology, Daegu Gyeongbuk Institute of Science and Technology, Daegu, Korea
2New Drug Development Center, Daegu-Gyeongbuk Medical Innovation Foundation, Daegu, Korea
- Corresponding author: Kyeong-Min Lee http://orcid.org/0000-0002-8031-7686 Division of Biotechnology, Daegu Gyeongbuk Institute of Science and Technology, 333 Techno jungang-daero, Hyeonpung-myeon, Dalseong-gun, Daegu 42988, Korea E-mail: leekm1009@dgist.ac.kr
- *Kyeong-Min Lee and Yeo Jin Hwang contributed equally to this study as first authors.
Copyright © 2024 Korean Diabetes Association
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
ABSTRACT
-
Background
- Renal fibrosis is characterized by the accumulation of extracellular matrix proteins and interstitial fibrosis. Alantolactone is known to exert anticancer, anti-inflammatory, antimicrobial and antifungal effects; however, its effects on renal fibrosis remains unknown. Here, we investigated whether alantolactone attenuates renal fibrosis in mice unilateral ureteral obstruction (UUO) and evaluated the effect of alantolactone on transforming growth factor (TGF) signaling pathway in renal cells.
-
Methods
- To evaluate the therapeutic effect of alantolactone, cell counting kit-8 (CCK-8) assay, histological staining, Western blot analysis, and real-time quantitative polymerase chain reaction were performed in UUO kidneys in vivo and in TGF-β-treated renal cells in vitro.
-
Results
- Alantolactone (0.25 to 4 µM) did not affect the viability of renal cells. Mice orally administered 5 mg/kg of alantolactone daily for 15 days did not show mortality or liver toxicity. Alantolactone decreased UUO-induced blood urea nitrogen and serum creatinine levels. In addition, it significantly alleviated renal tubulointerstitial damage and fibrosis and decreased collagen type I, fibronectin, and α-smooth muscle actin (α-SMA) expression in UUO kidneys. In NRK-49F cells, alantolactone inhibited TGF-β-stimulated expression of fibronectin, collagen type I, plasminogen activator inhibitor-1 (PAI-1), and α-SMA. In HK-2 cells, alantolactone inhibited TGF-β-stimulated expression of collagen type I and PAI-1. Alantolactone inhibited UUO-induced phosphorylation of Smad3 in UUO kidneys. In addition, it not only decreased TGF-β secretion but also Smad3 phosphorylation and translocation to nucleus in both kidney cell lines.
-
Conclusion
- Alantolactone improves renal fibrosis by inhibiting the TGF-β/Smad3 signaling pathway in obstructive nephropathy. Thus, alantolactone is a potential therapeutic agent for chronic kidney disease.
- Renal fibrosis is the final progressive manifestation of chronic kidney disease leading to end-stage renal disease, it is characterized by the activation of interstitial myofibroblasts and excessive accumulation of extracellular matrix (ECM) proteins. Transforming growth factor β (TGF-β) plays a crucial role all these processes [1-5].
- TGF-β expression is upregulated in various experimental and human renal diseases, where it induces the production of ECM proteins, including collagen type I and fibronectin, and renal tissue fibrosis [6-8]. TGF-β interacts with TGF-β receptors and activates intercellular mediators, known as Smad3 [5,9]. Phosphorylation of Smad3 by TGF-β has been observed in diabetic nephropathy [10,11], obstructive nephropathy [12], and drug toxicity-related nephropathy [13,14]. Smad3 mediates TGF-β-induced ECM deposition and renal fibrosis [5].
- Alantolactone (AL) is extracted from Inula species, which is traditionally used as a Chinese herbal medicine against allergic contact dermatitis [15], and is an effective an anti-tumoral [16], anti-inflammatory [17], and antioxidant agent [18]. Previous studies have reported its anti-inflammatory effects on the kidneys in mice with streptozotocin-induced diabetes [19], protective effects against neuronal cell death and amnesia in mice [20], and anti-fibrotic effects in pulmonary fibrosis [21]. However, its functional role in renal fibrosis has not been investigated in vivo and in vitro.
- In this study, we investigated the effect of AL on Smad3 inhibition and acceleration of renal interstitial fibrosis in unilateral ureteral obstruction (UUO)-induced renal fibrosis.
INTRODUCTION
- Animals and UUO model
- Male C57BL/6 8-week-old mice, weighing 25±2 g, were purchased from the Koatech Technology Corporation (Pyeongtaek, Korea). All experiments were approved by the Animal Care and Use Committee of Daegu Gyeongbuk Institute of Science and Technology (DGIST-IACUC-19052105-01). Mice were bred under standard room temperature (22°C±2°C) and humidity (60%±10%) conditions with a 12-hour light/dark cycle. Animals were randomly divided into the following three groups: control (CON, n=7), UUO (n=7), UUO with 5 mg/kg AL treatment (UUO/AL, n=7). AL (Carbosynth, Compton, UK) was dissolved in a filtered solution (1.25% dimethyl sulfoxide [DMSO], 2% polyethylene glycol [PEG] 400, 0.5% Tween 20 in saline), which was administered orally by gavage in equivalent volumes to CON and UUO mice once per day for 15 days, including 5 days of pretreatment. The UUO surgery was performed as previously described [22]: briefly, anesthetized mice underwent laparotomy to ligate the left with sterilized 5–0 silk for 10 days. Fibrosis-induced kidneys were fixed in a 10% formalin solution (Sigma-Aldrich, Burlington, MA, USA) or stored in liquid nitrogen for use as experimental samples.
- Blood analysis
- The serum of mice, separated using centrifugation of blood, was used to measure the levels of serum creatinine (Scr), blood urea nitrogen (BUN), alanine aminotransferase and aspartate aminotransferase using an automatic biochemical analyzer (Cobas Integra 800, Roche, Basel, Switzerland).
- Cell culture
- Human proximal tubule cell line (HK-2) and rat fibroblast cell line (NRK-49F) were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). HK-2 cells were grown in keratinocyte serum-free medium (Gibco, Waltham, MA, USA), supplemented with 50 µg/mL bovine pituitary extract and 5 ng/mL human recombinant epidermal growth factor. NRK-49F cells were cultured in Dulbecco’s modified Eagle medium/nutrient mixture F-12 medium (Gibco) with 5% fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin (Welgene, Gyeongsan, Korea). Cells were cultured in an atmosphere of 5% CO2 at 37°C in a humidity chamber and treated with 5 and 2 ng/mL of TGF-β1 (R&D Systems, Minneapolis, MN, USA), respectively, for the indicated time periods.
- Western blot analysis
- Total protein from cells and kidney tissues were obtained by homogenization in radioimmunoprecipitation assay (RIPA) lysis buffer (Thermo Fisher Scientific, Waltham, MA, USA) containing cOmplete, Mini Protease Inhibitor Cocktail (Roche), and Halt Phosphatase Inhibitor Cocktail (Thermo Fisher Scientific). Samples with an equal protein concentration were electrophoresed on Mini-PROTEAN Gels (Bio-Rad, Hercules, CA, USA), and then electrotransferred onto polyvinylidene fluoride membranes (Bio-Rad). After blocking with 5% bovine serum albumin (Sigma-Aldrich) for 1 hour, the membranes were immunoblotted overnight at 4°C with one of the following primary antibodies: anti-collagen type I (1:500) and anti-fibronectin (1:1,000) from Abcam (Cambridge, UK); anti-vimentin (1:1,000), anti-α-smooth muscle actin (α-SMA; 1:1,000), anti-plasminogen activator inhibitor-1 (PAI-1; 1:500) from BD Biosciences (Franklin Lakes, NJ, USA); and anti-phospho-Smad3 (1:500), anti-phospho-signal transducer and activator of transcription 3 (STAT3; 1: 1,000), anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH; 1:1,000), and anti-β-actin (1:1,000) from Santa Cruz Biotechnology (Dallas, TX, USA). The membranes were subsequently treated horseradish peroxidase (HRP)-linked secondary antibody (1:3,500). Protein expression was measured by visualizing the proteins bands using ChemiDoc XRS+ (Bio-Rad) and performing densitometric analysis using the Image J program 1.53e (National Institutes of Health, Bethesda, MD, USA).
- mRNA expression analysis using real time-quantitative polymerase chain reaction
- An RNA Extraction Kits (Takara, Tokyo, Japan) was used to extract total RNA. cDNA was synthesized using a Reverse Transcription Kit (Applied Biosystems, Waltham, MA, USA) and, mRNA levels were detected using an SYBR Green polymerase chain reaction (PCR) Master Mix kit (Applied Biosystems) on an ABI 7500 quantitative polymerase chain reaction (qPCR) system. The qPCR conditions and primer sequences were designed using the Primer Express Software ver. 3.0 (Thermo Fisher Scientific). The sequences of all the primers are provided in Table 1.
- Histological analysis and immunohistochemical staining
- Sirius red and Masson’s trichrome (Sigma-Aldrich) and immunohistochemical staining (Thermo Fisher Scientific) were performed on paraffin sections (4 μm thick) according to the manufacturer’s instructions. Kidney sections were deparaffinized using xylene and rehydrated using graded ethanol series. Tubulointerstitial collagen deposition was assessed using Sirius red or Masson’s trichrome staining. For Sirius red staining, the slides were immersed for 18 hours in saturated picric acid with 0.1% Sirius red F3BA, and 0.01N hydrochloric acid for 2 minutes. For Masson’s trichrome staining, slides were treated with Bouin’s solution and incubated sequentially with hematoxylin for 10 minutes, Biebrich scarlet-acid fuchsin for 5 minutes, phosphotungstic acid/phosphomolybdic acid for 10 minutes, aniline blue for 15 minutes, and 1% acetic acid for 5 minutes. Immunohistochemical staining was performed with anti-collagen type I (1:200), anti-fibronectin (1:200, Abcam), anti-α-SMA (1:300), anti-p-Smad3 (1:200, Santa Cruz Biotechnology), and DAB staining kit (Roche). The kidney sections were dehydrated with a graded series of ethanol and xylene and mounted on glass slides for microscopic investigation. After staining, sections from each kidney of five different animals were selected and the images (×200) were taken using Leica Microscope (Leica Microsystem, Wetzlar, Germany) and Leica Application Suite V3.8 software (Leica Microsystems). Quantification of the aniline blue (collagen, blue), Sirius red (collagen fiber, red), and immunostaining (brown color) positive areas was performed using computer-based morphometric analysis.
- Immunofluorescence staining
- HK-2 and NRK-49F cells, seeded on 8-well glass slides (Millipore, Burlington, MA, USA) were treated with AL or TGF-β1. For fixation, the glass slides were incubated with 2% paraformaldehyde for 2 hours at room temperature (20°C to 25°C). After washing the cells with phosphate-buffered saline (PBS), they were permeabilized for 2 minutes using 0.3% Triton X-100 (Sigma-Aldrich) in PBS. Cells were blocked with CAS-Block Histochemical Reagent (Thermo Fisher Scientific) for 1 hour, and then incubated overnight with anti-p-Smad3 antibody (1:100, Abcam) at room temperature. The slides were washed three times for 5 minutes each with PBS and then incubated with Alexa Fluor 488 anti-rabbit antibody for 1 hour in the dark. After staining the nuclei with 4’,6-diamidino-2-phenylindole, dihydrochloride (DAPI, Invitrogen) for 5 minutes, the cells were washed three times with distilled water prior to mounting with a fluorescent mounting medium (Sigma-Aldrich). Immunofluorescence was detected using a laser scanning confocal microscope (Zeiss, Oberkochen, Baden-Württemberg, Germany) and analyzed using the ZEN 2.6 (blue edition) software.
- Statistical analysis
- Data are expressed as the mean±standard error of the mean of three independent experiments. Statistical analysis was performed using Student’s t-test. Statistical significance was set at P<0.05.
METHODS
- Toxicity of alantolactone
- To examine the cell toxicity of AL, NRK-49F and HK-2 cells were incubated with the indicated concentrations of AL and their viability was measured using the cell counting kit-8 (CCK-8) assay. AL did not affect the cell viability at 0.25 to 4 µM but at 10 µM the viability of both the renal cell lines was less than 90% (Fig. 1A and B). Therefore, we used 1, 2, and 4 µM of AL for the subsequent in vitro experiments. In vivo toxicity of AL, was assessed in mice orally administered 5 or 10 mg/kg of AL daily for 15 days; these mice did not exhibit mortality, any unusual behavior, and liver toxicity (Supplementary Fig. 1).
- Alantolactone attenuate UUO-induced renal dysfunction and renal fibrosis
- As shown in Fig. 1C and D, the levels of Scr and BUN were increased in the UUO group. Administration of AL in UUO mice led to lower Scr and BUN levels compared with those in the UUO mice that were not treated.
- A comparison of results for the UUO and UUO/AL groups (Fig. 2A-C) showed that UUO kidneys exhibited significantly increased tubular atrophy and tubulointerstitial fibrosis. Conversely, UUO-induced renal fibrosis was markedly decreased in the kidneys of AL-treated mice. Masson’s trichrome and Sirius red staining showed that interstitial collagen deposition in UUO kidneys increased significantly, whereas less intense staining was observed in the kidneys of AL-treated mice. Immunohistochemical staining showed that the expression of collagen type I, fibronectin, and α-SMA was increased in UUO kidneys. In contrast, the expression of all the fibrotic factors was decreased in AL-treated UUO kidneys (Fig. 2A and D-F). Similar results were also observed in immunoblot analysis of kidney tissue lysates. The expression of these fibrotic factors proteins was significantly increased in UUO kidneys and decreased in AL-treated UUO kidneys (Fig. 2G-J).
- Alantolactone inhibits the expression of TGF-β-induced renal fibrotic factors in renal fibroblasts and renal proximal tubule cells
- Next, the effects of AL on TGF-β-stimulated profibrotic target gene inhibition and on protein expression in NRK-49F and HK-2 cells were evaluated. Real-time qPCR analysis showed that AL dose-dependently inhibited the expression of TGF-β-stimulated collagen type I, fibronectin, PAI-1, and α-SMA mRNAs in NRK-49F cells (Fig. 3A-D). In addition, AL effectively inhibited the expression of TGF-β-stimulated profibrotic factor proteins in NRK-49F cells (Fig. 3E). AL also inhibited TGF-β-stimulated expreesion of collagen type I and PAI-1 mRNA and protein in HK-2 cells (Fig. 3F-H).
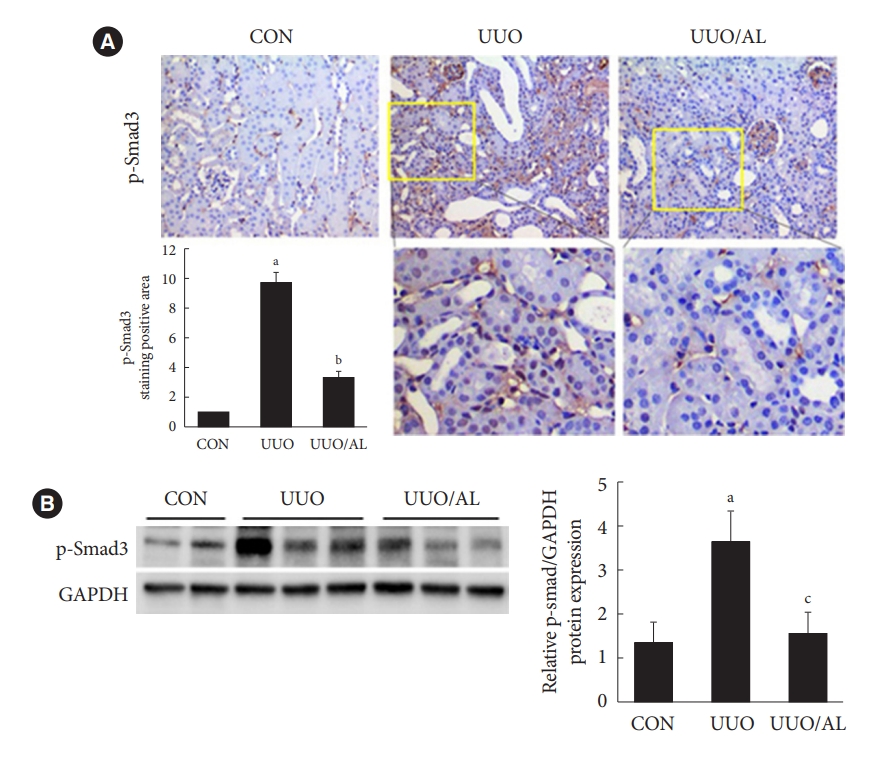
- Alantolactone ameliorates renal fibrosis by inhibition of Smad3 phosphorylation in UUO kidneys and renal cells
- TGF-β regulates renal fibrotic genes and matrix proteins through Smad signaling, and subsequently contributes to renal fibroblast activation and tubulointerstitial fibrosis. To elucidate the inhibitory mechanism of renal fibrosis, we investigated whether AL regulated Smad3, which is activated in UUO kidneys and TGF-β-treated renal cells. In the UUO kidneys, phospho-Smad3 levels increased compared with those in CON kidneys and were significantly decreased in AL-treated UUO kidneys (Fig. 4A). In addition, the protein expression in UUO kidneys was significantly inhibited by AL administration (Fig. 4B). As shown in Fig. 5A and B, AL inhibited TGF-β secretion in cultured renal fibroblasts and tubular epithelium-like cells. Immunoblot analysis showed that AL inhibited TGF-β-stimulated Smad3 phosphorylation as well as Smad3 translocation (Fig. 5E) to nucleus in both the renal cells (Fig. 5C and D).
RESULTS
- This study is the first to demonstrate that AL, a natural compound produced by Inula helenium, effectively ameliorated renal fibrosis during obstructive nephropathy. AL prevented the expression of renal interstitial fibrotic factors including type I collagen, fibronectin, α-SMA, and PAI-1 in UUO kidneys and TGF-β-treated renal cells. This inhibitory effect of AL on renal fibrosis proceeds by first decreasing the secretion of TGF-β and thereby sequentially inhibiting phosphorylation and localization of Smad3.
- Pharmacokinetic studies have shown that the absorption of AL is via passive diffusion, and that its intestinal absorption is good [23]; the highest concentration of AL was achieved in the small intestine and feces clearance was shown to be the dominant elimination pathway for the lactones [24]. However, AL has a rapid onset and does not cause significant damage to normal animal tissues and organs [24-26]. Several studies have shown that AL exhibits multiple pharmacological activities [15-18]. We focused on the anti-fibrotic effect of AL in the study. To demonstrate the anti-fibrotic effects of AL on renal fibrosis, we used the UUO model with oral gavage administration. Zhu et al. [19] administered 5 or 10 mg/kg per 2 days AL by oral gavage for 8 weeks to confirm its protective effect on streptozotocin-induced renal inflammation. In several studies, the anticancer effects of AL were observed after administering AL at concentration ranging from 5 to 50 mg/kg to mouse or rats [25,27-29]. Based on the pharmacokinetic characteristics described in several reports and our preliminary studies, we selected 5 mg/kg of AL to demonstrate the anti-fibrotic effect in the UUO-induced renal fibrosis model.
- Kidneys with UUO present common features of obstructive nephropathy, such as tubular degeneration and atrophy, and tubulointerstitial fibrosis [22,30,31]. UUO kidneys at day 10 exhibited morphological change, such as tubulointerstitial injury and ECM deposition, accompanied by increased expression of fibrotic factors including collagen type I, fibronectin, and α-SMA. This in vivo study revealed that AL treatment preserved the tubular interstitial integrity and reduced ECM deposition in UUO kidneys. Furthermore, AL drastically decreased TGF-β-induced expression of collagen type I, fibronectin, PAI-1, and α-SMA in renal fibroblasts. AL also inhibited TGF-β-induced expression of collagen type I and PAI-1 in renal tubular cells.
- TGF-β has been reported to trigger the Smad signaling pathway related to tubulointerstitial fibrosis in experimental kidney fibrosis models [32] and to enhance the transcription of profibrotic genes, including those encoding α-SMA, PAI-1, and matrix proteins [33,34]. Our data demonstrate that AL drastically inhibited Smad3 phosphorylation in UUO-induced fibrotic kidneys and ameliorated renal fibrosis in vivo. In vitro, AL inhibited TGF-β-stimulated Smad3 phosphorylation and localization into the nucleus in renal cells as well as significantly reduced TGF-β secretion.
- AL is well known as a selective STAT3 inhibitor and its beneficial effects in many diseases, including glioblastoma, liver cancer, breast cancer, and inflammatory disease, have been focused upon in several studies [27,28,35,36]. We also confirmed that alantolactone inhibited TGF-β- or UUO-stimulated STAT3 phosphorylation in vivo and in vitro (Supplementary Fig. 2A and B). In addition, AL inhibited the expression of vimentin, an epithelial-mesenchymal transition (EMT) factor, and F4/80, a macrophage marker, in UUO kidneys (Supplementary Fig. 2C and D). These results show the potential of AL to inhibit EMT and inflammation, and studies on these aspects are ongoing. Qin et al. [37] showed that the potential of herb-drug interactions rises when they are co-administered with other drugs. Specifically, it has been suggested that AL might exhibit herb-drug interactions when co-administrated with drugs that are CYP3A4 and CYP2C19 substrates [37]. Clinical studies are warranted to further investigate the interactions of drugs co-administered with AL.
- The in vivo and in vitro results of the current study suggest that AL ameliorates renal fibrosis by regulating the TGF-β/Smad3 signaling pathway. This study is the first to demonstrate that AL inhibits renal fibrosis and the expression of its related factors in an animal model of obstructive nephropathy induced by UUO. In cultured renal cells, AL effectively inhibited the expression of renal fibrotic factors via downregulating TGF-β secretion, and Smad3 phosphorylation and translocation into the nucleus. Because oral administration of AL effectively attenuated renal fibrosis, it could be proposed as a therapeutic agent for fibrotic kidney disease. The use of AL for treatment of human kidney disease deserves further investigation.
DISCUSSION
SUPPLEMENTARY MATERIALS
Supplementary Fig. 1
Supplementary Fig. 2.
-
CONFLICTS OF INTEREST
No potential conflict of interest relevant to this article was reported.
-
AUTHOR CONTRIBUTIONS
Conception or design: K.M.L.
Acquisition, analysis, or interpretation of data: all authors.
Drafting the work or revising: all authors.
Final approval of the manuscript: K.M.L.
-
FUNDING
This work was supported by DGIST projects 21-BT-06, and NRF-2021R1A2C1011314 and NRF-2018R1C1B6008955 from the National Research Foundation of Korea funded by the Ministry of Science, ICT of the Republic of Korea.
NOTES
-
Acknowledgements
- None




- 1. Grgic I, Duffield JS, Humphreys BD. The origin of interstitial myofibroblasts in chronic kidney disease. Pediatr Nephrol 2012;27:183-93.ArticlePubMedPMCPDF
- 2. Liu Y. Cellular and molecular mechanisms of renal fibrosis. Nat Rev Nephrol 2011;7:684-96.ArticlePubMedPMCPDF
- 3. Zeisberg M, Neilson EG. Mechanisms of tubulointerstitial fibrosis. J Am Soc Nephrol 2010;21:1819-34.ArticlePubMed
- 4. Border WA, Okuda S, Languino LR, Ruoslahti E. Transforming growth factor-beta regulates production of proteoglycans by mesangial cells. Kidney Int 1990;37:689-95.PubMed
- 5. Meng XM, Tang PM, Li J, Lan HY. TGF-β/Smad signaling in renal fibrosis. Front Physiol 2015;6:82.ArticlePubMedPMC
- 6. Nakamura T, Miller D, Ruoslahti E, Border WA. Production of extracellular matrix by glomerular epithelial cells is regulated by transforming growth factor-beta 1. Kidney Int 1992;41:1213-21.PubMed
- 7. Jung GS, Hwang YJ, Choi JH, Lee KM. Lin28a attenuates TGF-β-induced renal fibrosis. BMB Rep 2020;53:594-9.ArticlePubMedPMC
- 8. Roberts AB, Heine UI, Flanders KC, Sporn MB. Transforming growth factor-beta: major role in regulation of extracellular matrix. Ann N Y Acad Sci 1990;580:225-32.PubMed
- 9. Miyazono K. TGF-beta signaling by Smad proteins. Cytokine Growth Factor Rev 2000;11:15-22.PubMed
- 10. Hills CE, Squires PE. The role of TGF-β and epithelial-to mesenchymal transition in diabetic nephropathy. Cytokine Growth Factor Rev 2011;22:131-9.ArticlePubMed
- 11. Li L, Emmett N, Mann D, Zhao X. Fenofibrate attenuates tubulointerstitial fibrosis and inflammation through suppression of nuclear factor-κB and transforming growth factor-β1/Smad3 in diabetic nephropathy. Exp Biol Med (Maywood) 2010;235:383-91.ArticlePubMedPMCPDF
- 12. Ai J, Nie J, He J, Guo Q, Li M, Lei Y, et al. GQ5 hinders renal fibrosis in obstructive nephropathy by selectively inhibiting TGF-β-induced Smad3 phosphorylation. J Am Soc Nephrol 2015;26:1827-38.ArticlePubMedPMC
- 13. Li J, Campanale NV, Liang RJ, Deane JA, Bertram JF, Ricardo SD. Inhibition of p38 mitogen-activated protein kinase and transforming growth factor-beta1/Smad signaling pathways modulates the development of fibrosis in adriamycin-induced nephropathy. Am J Pathol 2006;169:1527-40.PubMedPMC
- 14. Nam HK, Lee SJ, Kim MH, Rho JH, Son YK, Lee SM, et al. Rosuvastatin attenuates inflammation, apoptosis and fibrosis in a rat model of cyclosporine-induced nephropathy. Am J Nephrol 2013;37:7-15.ArticlePubMedPDF
- 15. Hoffmann D. Medical herbalism: the science and practice of herbal medicine. Fairfield: Healing Arts Press; 2003. p. 672.
- 16. Rasul A, Khan M, Ali M, Li J, Li X. Targeting apoptosis pathways in cancer with alantolactone and isoalantolactone. ScientificWorldJournal 2013;2013:248532.ArticlePubMedPMCPDF
- 17. Chun J, Choi RJ, Khan S, Lee DS, Kim YC, Nam YJ, et al. Alantolactone suppresses inducible nitric oxide synthase and cyclooxygenase-2 expression by down-regulating NF-κB, MAPK and AP-1 via the MyD88 signaling pathway in LPS-activated RAW 264.7 cells. Int Immunopharmacol 2012;14:375-83.ArticlePubMed
- 18. Seo JY, Lim SS, Kim JR, Lim JS, Ha YR, Lee IA, et al. Nrf2-mediated induction of detoxifying enzymes by alantolactone present in Inula helenium. Phytother Res 2008;22:1500-5.ArticlePubMed
- 19. Zhu Y, Ling Y, Wang X. Alantolactone mitigates renal injury induced by diabetes via inhibition of high glucose-mediated inflammatory response and macrophage infiltration. Immunopharmacol Immunotoxicol 2020;42:84-92.ArticlePubMed
- 20. Seo JY, Lim SS, Kim J, Lee KW, Kim JS. Alantolactone and isoalantolactone prevent amyloid β25-35-induced toxicity in mouse cortical neurons and scopolamine-induced cognitive impairment in mice. Phytother Res 2017;31:801-11.ArticlePubMedPDF
- 21. Zhao WY, Luan ZL, Liu TT, Ming WH, Huo XK, Huang HL, et al. Inula japonica ameliorated bleomycin-induced pulmonary fibrosis via inhibiting soluble epoxide hydrolase. Bioorg Chem 2020;102:104065.ArticlePubMed
- 22. Dai Y, Zhang W, Wen J, Zhang Y, Kellems RE, Xia Y. A2B adenosine receptor-mediated induction of IL-6 promotes CKD. J Am Soc Nephrol 2011;22:890-901.ArticlePubMedPMC
- 23. Xu R, Peng Y, Wang M, Li X. Intestinal absorption of isoalantolactone and alantolactone, two sesquiterpene lactones from Radix Inulae, using Caco-2 cells. Eur J Drug Metab Pharmacokinet 2019;44:295-303.ArticlePubMedPDF
- 24. Xu R, Zhou G, Peng Y, Wang M, Li X. Pharmacokinetics, tissue distribution and excretion of isoalantolactone and alantolactone in rats after oral administration of Radix Inulae extract. Molecules 2015;20:7719-36.ArticlePubMedPMC
- 25. Lee JY, Kim SB, Chun J, Song KH, Kim YS, Chung SJ, et al. High body clearance and low oral bioavailability of alantolactone, isolated from Inula helenium, in rats: extensive hepatic metabolism and low stability in gastrointestinal fluids. Biopharm Drug Dispos 2016;37:156-67.PubMed
- 26. Jiang Y, Xu H, Wang J. Alantolactone induces apoptosis of human cervical cancer cells via reactive oxygen species generation, glutathione depletion and inhibition of the Bcl-2/Bax signaling pathway. Oncol Lett 2016;11:4203-7.ArticlePubMedPMC
- 27. Wang X, Yu Z, Wang C, Cheng W, Tian X, Huo X, et al. Alantolactone, a natural sesquiterpene lactone, has potent antitumor activity against glioblastoma by targeting IKKβ kinase activity and interrupting NF-κB/COX-2-mediated signaling cascades. J Exp Clin Cancer Res 2017;36:93.ArticlePubMedPMCPDF
- 28. Liu YR, Cai QY, Gao YG, Luan X, Guan YY, Lu Q, et al. Alantolactone, a sesquiterpene lactone, inhibits breast cancer growth by antiangiogenic activity via blocking VEGFR2 signaling. Phytother Res 2018;32:643-50.ArticlePubMedPDF
- 29. Yin C, Dai X, Huang X, Zhu W, Chen X, Zhou Q, et al. Alantolactone promotes ER stress-mediated apoptosis by inhibition of TrxR1 in triple-negative breast cancer cell lines and in a mouse model. J Cell Mol Med 2019;23:2194-206.ArticlePubMedPMCPDF
- 30. Chevalier RL, Forbes MS, Thornhill BA. Ureteral obstruction as a model of renal interstitial fibrosis and obstructive nephropathy. Kidney Int 2009;75:1145-52.ArticlePubMed
- 31. Tashiro K, Tamada S, Kuwabara N, Komiya T, Takekida K, Asai T, et al. Attenuation of renal fibrosis by proteasome inhibition in rat obstructive nephropathy: possible role of nuclear factor kappaB. Int J Mol Med 2003;12:587-92.PubMed
- 32. Sato M, Muragaki Y, Saika S, Roberts AB, Ooshima A. Targeted disruption of TGF-beta1/Smad3 signaling protects against renal tubulointerstitial fibrosis induced by unilateral ureteral obstruction. J Clin Invest 2003;112:1486-94.ArticlePubMedPMC
- 33. Meng XM, Huang XR, Xiao J, Chen HY, Zhong X, Chung AC, et al. Diverse roles of TGF-β receptor II in renal fibrosis and inflammation in vivo and in vitro. J Pathol 2012;227:175-88.ArticlePubMed
- 34. Samarakoon R, Overstreet JM, Higgins SP, Higgins PJ. TGF-β1 → SMAD/p53/USF2 → PAI-1 transcriptional axis in ureteral obstruction-induced renal fibrosis. Cell Tissue Res 2012;347:117-28.ArticlePubMedPMCPDF
- 35. Khan M, Li T, Ahmad Khan MK, Rasul A, Nawaz F, Sun M, et al. Alantolactone induces apoptosis in HepG2 cells through GSH depletion, inhibition of STAT3 activation, and mitochondrial dysfunction. Biomed Res Int 2013;2013:719858.ArticlePubMedPMCPDF
- 36. Park JY, Yoo KD, Bae E, Kim KH, Lee JW, Shin SJ, et al. Blockade of STAT3 signaling alleviates the progression of acute kidney injury to chronic kidney disease through antiapoptosis. Am J Physiol Renal Physiol 2022;322:F553-72.ArticlePubMed
- 37. Qin CZ, Lv QL, Wu NY, Cheng L, Chu YC, Chu TY, et al. Mechanism-based inhibition of alantolactone on human cytochrome P450 3A4 in vitro and activity of hepatic cytochrome P450 in mice. J Ethnopharmacol 2015;168:146-9.ArticlePubMed
 PubReader
PubReader ePub Link
ePub Link Cite
Cite