- Current
- Browse
- Collections
-
For contributors
- For Authors
- Instructions to authors
- Article processing charge
- e-submission
- For Reviewers
- Instructions for reviewers
- How to become a reviewer
- Best reviewers
- For Readers
- Readership
- Subscription
- Permission guidelines
- About
- Editorial policy
Articles
- Page Path
- HOME > Diabetes Metab J > Volume 37(5); 2013 > Article
-
ReviewPathophysiology Regulation of Muscle Pyruvate Dehydrogenase Complex in Insulin Resistance: Effects of Exercise and Dichloroacetate
- Dumitru Constantin-Teodosiu
-
Diabetes & Metabolism Journal 2013;37(5):301-314.
DOI: https://doi.org/10.4093/dmj.2013.37.5.301
Published online: October 17, 2013
- 6,325 Views
- 80 Download
- 47 Crossref
School of Life Sciences, University of Nottingham Medical School, Nottingham, UK.
- Corresponding author: Dumitru Constantin-Teodosiu. School of Life Sciences, University of Nottingham Medical School, NG7 2UH, Nottingham, UK. tim.constantin@nottingham.ac.uk
Copyright © 2013 Korean Diabetes Association
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
- ABSTRACT
- INTRODUCTION
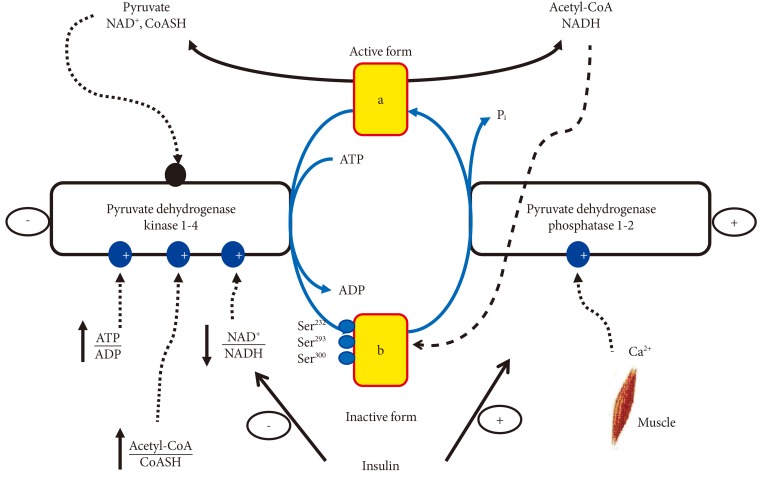
- REGULATION OF PDC ACTIVITY
- MUSCLE PDC ACTIVITY AND MUSCLE ACETYLCARNITINE ACCUMULATION
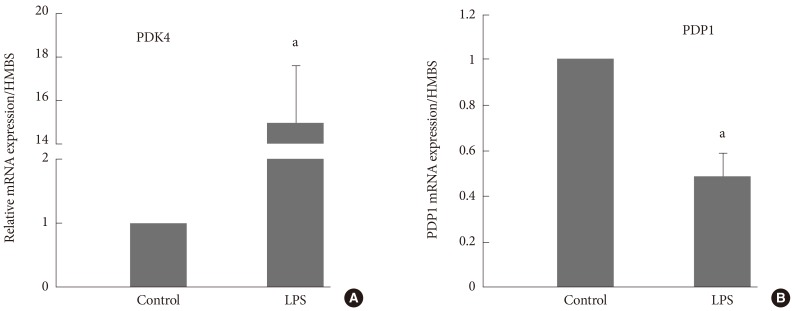
- PDC AND PDK4 IN HUMAN SKELETAL MUSCLE
- INHIBITION OF MUSCLE PDC ACTIVITY BY DRUGS
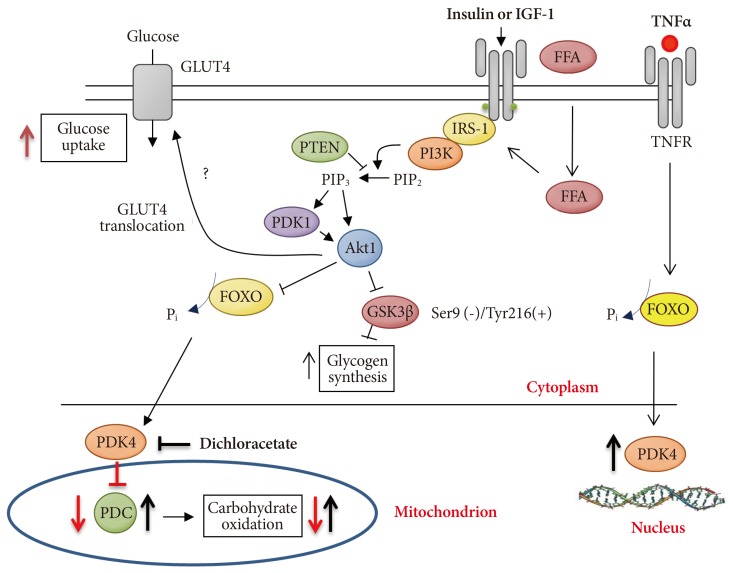
- MECHANISM OF PDK4 ACTIVATION BY PPARs/FOXO1
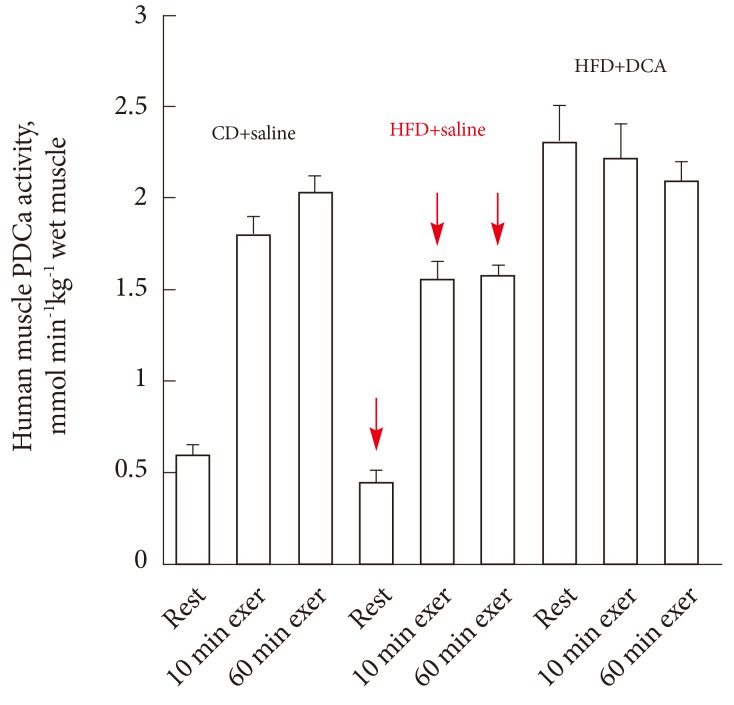
- EFFECT OF DICHLOROACETATE ON PDC ACTIVITY AND CHO OXIDATION
- DOES THE HYPOXIA-INDUCIBLE FACTOR-1α MEDIATED PDK4 UP-REGULATION MAKE CHUVASH POLYCYTHEMIA PATIENTS INSULIN RESISTANT?
- CONCLUSIONS
- NOTES
- REFERENCES


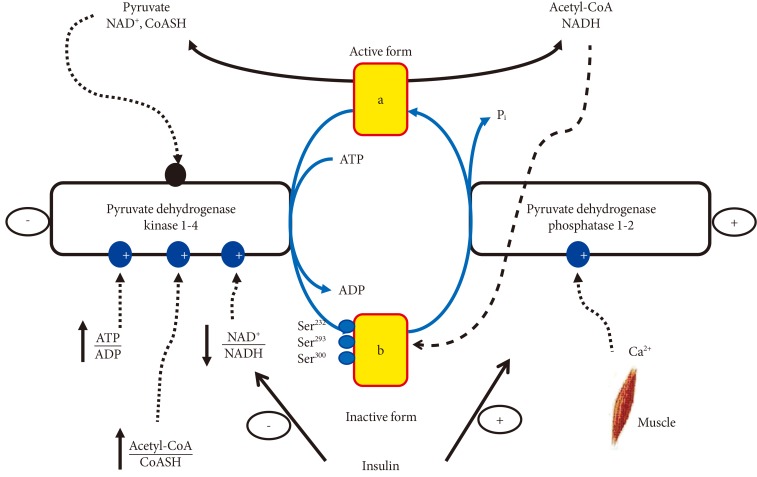
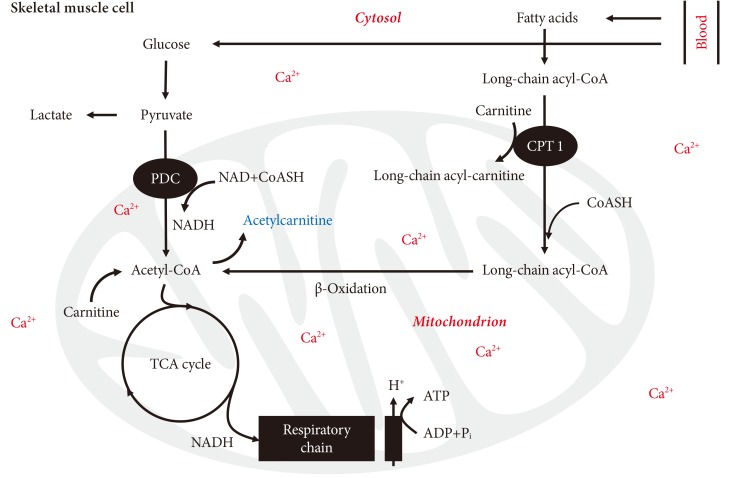
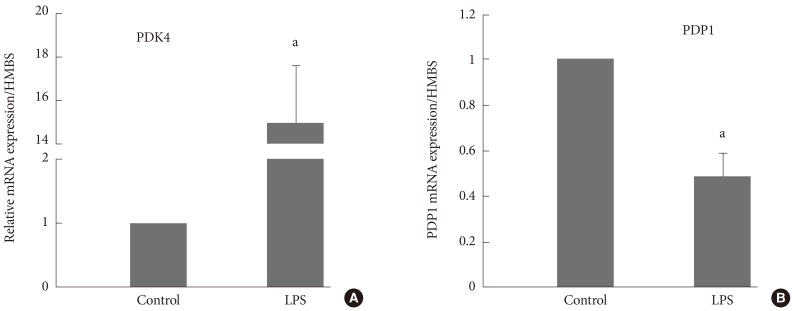
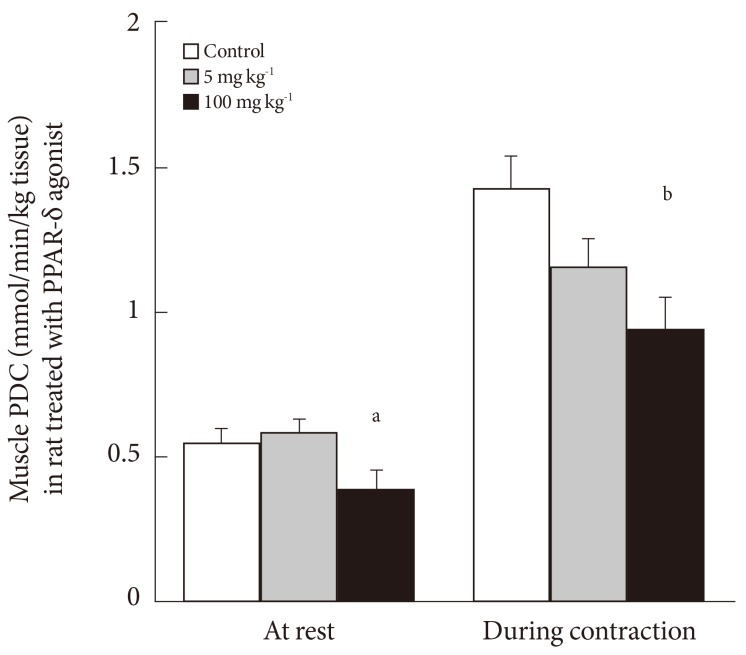


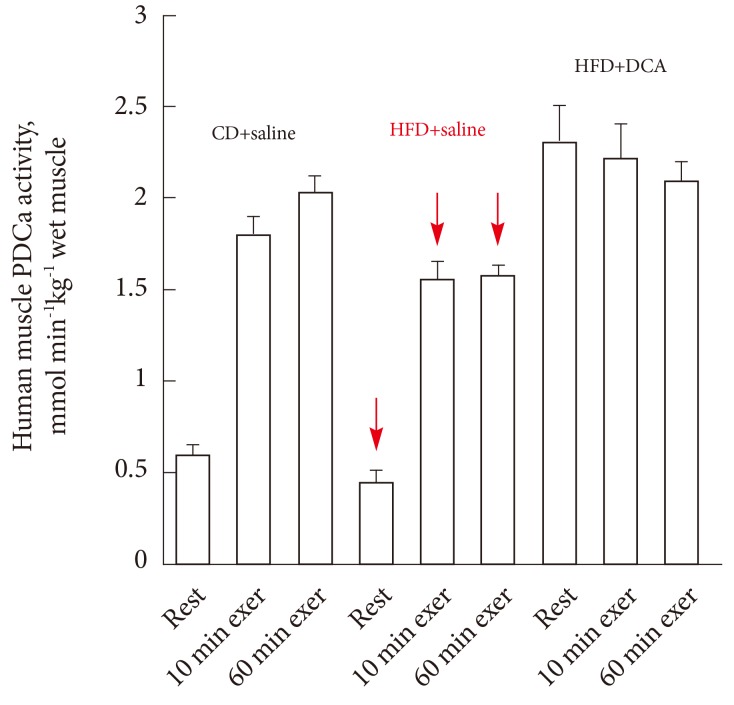
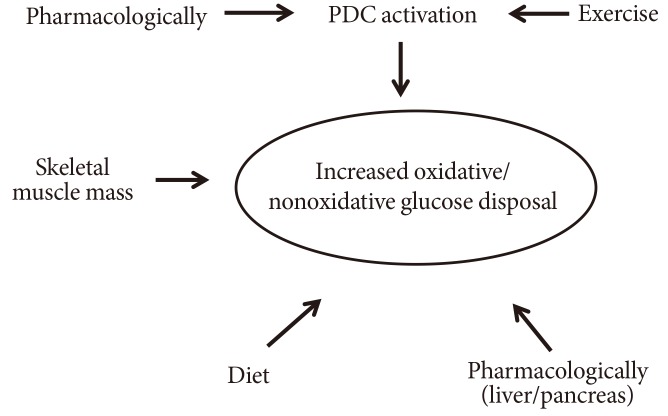
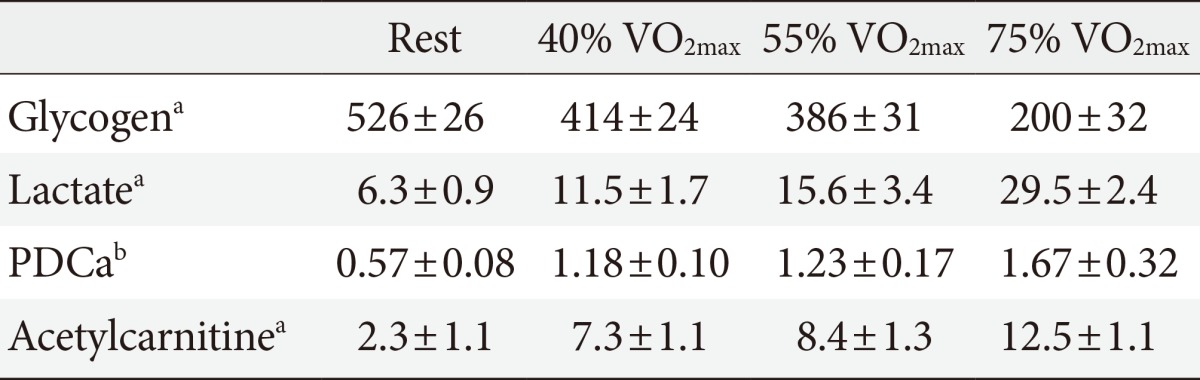
Figure & Data
References
Citations
Citations to this article as recorded by 
- Oxidative stress and metabolism meet epigenetic modulation in physical exercise
José Luis García-Giménez, Irene Cánovas-Cervera, Federico V. Pallardó
Free Radical Biology and Medicine.2024; 213: 123. CrossRef - The pyruvate dehydrogenase complex: Life’s essential, vulnerable and druggable energy homeostat
Peter W. Stacpoole, Charles E. McCall
Mitochondrion.2023; 70: 59. CrossRef - Regulating mitochondrial metabolism by targeting pyruvate dehydrogenase with dichloroacetate, a metabolic messenger
Nick Schoenmann, Nicholas Tannenbaum, Ryan M. Hodgeman, Raghavan Pillai Raju
Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease.2023; 1869(7): 166769. CrossRef - Serum metabolomics profiling by proton nuclear magnetic resonance spectrometry of the response to single oral macronutrient challenges in women with polycystic ovary syndrome (PCOS) compared with male and female controls
Héctor F. Escobar-Morreale, María Ángeles Martínez-García, María Insenser, Nicolau Cañellas, Xavier Correig, Manuel Luque-Ramírez
Biology of Sex Differences.2023;[Epub] CrossRef - Metabolomics and mitochondrial dysfunction in cardiometabolic disease
Abhishek Shastry, Kimberly Dunham-Snary
Life Sciences.2023; 333: 122137. CrossRef - Loss of metabolic flexibility as a result of overexpression of pyruvate dehydrogenase kinases in muscle, liver and the immune system: Therapeutic targets in metabolic diseases
Jae‐Han Jeon, Themis Thoudam, Eun Jung Choi, Min‐Ji Kim, Robert A Harris, In‐Kyu Lee
Journal of Diabetes Investigation.2021; 12(1): 21. CrossRef - Targeting pyruvate dehydrogenase kinase signaling in the development of effective cancer therapy
Saleha Anwar, Anas Shamsi, Taj Mohammad, Asimul Islam, Md. Imtaiyaz Hassan
Biochimica et Biophysica Acta (BBA) - Reviews on Cancer.2021; 1876(1): 188568. CrossRef - Effect of exercise training on skeletal muscle protein expression in relation to insulin sensitivity: Per‐protocol analysis of a randomized controlled trial (GO‐ACTIWE)
Lea Bruhn, Rasmus Kjøbsted, Jonas Salling Quist, Anne Sofie Gram, Mads Rosenkilde, Kristine Færch, Jørgen F.P. Wojtaszewski, Bente Stallknecht, Martin Bæk Blond
Physiological Reports.2021;[Epub] CrossRef - The Mechanism behind Influenza Virus Cytokine Storm
Yinuo Gu, Xu Zuo, Siyu Zhang, Zhuoer Ouyang, Shengyu Jiang, Fang Wang, Guoqiang Wang
Viruses.2021; 13(7): 1362. CrossRef - Suppression of Pyruvate Dehydrogenase Kinase by Dichloroacetate in Cancer and Skeletal Muscle Cells Is Isoform Specific and Partially Independent of HIF-1α
Nives Škorja Milić, Klemen Dolinar, Katarina Miš, Urška Matkovič, Maruša Bizjak, Mojca Pavlin, Matej Podbregar, Sergej Pirkmajer
International Journal of Molecular Sciences.2021; 22(16): 8610. CrossRef - The Regulatory Roles of PPARs in Skeletal Muscle Fuel Metabolism and Inflammation: Impact of PPAR Agonism on Muscle in Chronic Disease, Contraction and Sepsis
Hannah Crossland, Dumitru Constantin-Teodosiu, Paul L. Greenhaff
International Journal of Molecular Sciences.2021; 22(18): 9775. CrossRef - PPARα, δ and FOXO1 Gene Silencing Overturns Palmitate-Induced Inhibition of Pyruvate Oxidation Differentially in C2C12 Myotubes
Hung-Che Chien, Despina Constantin, Paul L. Greenhaff, Dumitru Constantin-Teodosiu
Biology.2021; 10(11): 1098. CrossRef - Molecular Mechanisms of Muscle Fatigue
Dumitru Constantin-Teodosiu, Despina Constantin
International Journal of Molecular Sciences.2021; 22(21): 11587. CrossRef - Coronavirus Disease-19 (COVID-19) and Modern Lifestyle Diseases
Pallav Sengupta, Sulagna Dutta
Biomedical and Pharmacology Journal.2021; 14(4): 2245. CrossRef - A single bout of resistance exercise improves postprandial lipid metabolism in overweight/obese men with prediabetes
Adam J. Bittel, Daniel C. Bittel, Bettina Mittendorfer, Bruce W. Patterson, Adewole L. Okunade, Jun Yoshino, Lane C. Porter, Nada A. Abumrad, Dominic N. Reeds, W. Todd Cade
Diabetologia.2020; 63(3): 611. CrossRef - An integrative approach to the regulation of mitochondrial respiration during exercise: Focus on high-intensity exercise
Jose A.L. Calbet, Saúl Martín-Rodríguez, Marcos Martin-Rincon, David Morales-Alamo
Redox Biology.2020; 35: 101478. CrossRef - PPARδ and FOXO1 Mediate Palmitate-Induced Inhibition of Muscle Pyruvate Dehydrogenase Complex and CHO Oxidation, Events Reversed by Electrical Pulse Stimulation
Hung-Che Chien, Paul L. Greenhaff, Dumitru Constantin-Teodosiu
International Journal of Molecular Sciences.2020; 21(16): 5942. CrossRef - Cancer cachexia has many symptoms but only one cause: anoxia
Tomas Koltai
F1000Research.2020; 9: 250. CrossRef - Structural basis for the inhibition of PDK2 by novel ATP- and lipoyl-binding site targeting compounds
Jihoon Kang, Haushabhau S. Pagire, Donguk Kang, Yo Han Song, In Kyu Lee, Kang Taek Lee, Chin-Ju Park, Jin Hee Ahn, Jungwook Kim
Biochemical and Biophysical Research Communications.2020; 527(3): 778. CrossRef - Influenza and obesity: its odd relationship and the lessons for COVID-19 pandemic
Livio Luzi, Maria Grazia Radaelli
Acta Diabetologica.2020; 57(6): 759. CrossRef - Changes in gene expression of lactate carriers (MCT1 and CD147) in cardiac muscle of diabetic male rats: the effect of dichloroacetate and endurance training
H. Rezaeinasab, A. Habibi, M. Nikbakht, M. Rashno, S. Shakerian
The Ukrainian Biochemical Journal.2020; 92(5): 111. CrossRef - Mitochondrial Uncoupling Coordinated With PDH Activation Safely Ameliorates Hyperglycemia via Promoting Glucose Oxidation
Haowen Jiang, Jia Jin, Yanan Duan, Zhifu Xie, Yufeng Li, Anhui Gao, Min Gu, Xinwen Zhang, Chang Peng, Chunmei Xia, Tiancheng Dong, Hui Li, Lifang Yu, Jie Tang, Fan Yang, Jingya Li, Jia Li
Diabetes.2019; 68(12): 2197. CrossRef - Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Induced by Repeated Forced Swimming in Mice
Takuya Ohba, Shinichi Domoto, Miyu Tanaka, Shinsuke Nakamura, Masamitsu Shimazawa, Hideaki Hara
Biological and Pharmaceutical Bulletin.2019; 42(7): 1140. CrossRef - The Beta Cell in Type 2 Diabetes
Ashley A. Christensen, Maureen Gannon
Current Diabetes Reports.2019;[Epub] CrossRef - Reduced expression of Twist 1 is protective against insulin resistance of adipocytes and involves mitochondrial dysfunction
Sumei Lu, Hong Wang, Rui Ren, Xiaohong Shi, Yanmei Zhang, Wanshan Ma
Scientific Reports.2018;[Epub] CrossRef - PDK4 Deficiency Suppresses Hepatic Glucagon Signaling by Decreasing cAMP Levels
Bo-Yoon Park, Jae-Han Jeon, Younghoon Go, Hye Jin Ham, Jeong-Eun Kim, Eun Kyung Yoo, Woong Hee Kwon, Nam-Ho Jeoung, Yong Hyun Jeon, Seung-Hoi Koo, Byung-Gyu Kim, Ling He, Keun-Gyu Park, Robert A. Harris, In-Kyu Lee
Diabetes.2018; 67(10): 2054. CrossRef - Differences in Muscle Metabolism Between Triathletes and Normally Active Volunteers Investigated Using Multinuclear Magnetic Resonance Spectroscopy at 7T
Radka Klepochová, Ladislav Valkovič, Thomas Hochwartner, Christoph Triska, Norbert Bachl, Harald Tschan, Siegfried Trattnig, Michael Krebs, Martin Krššák
Frontiers in Physiology.2018;[Epub] CrossRef - Defining the contribution of skeletal muscle pyruvate dehydrogenase α1 to exercise performance and insulin action
Kristoffer Svensson, Jessica R. Dent, Shahriar Tahvilian, Vitor F. Martins, Abha Sathe, Julien Ochala, Mulchand S. Patel, Simon Schenk
American Journal of Physiology-Endocrinology and Metabolism.2018; 315(5): E1034. CrossRef - Early-onset and classical forms of type 2 diabetes show impaired expression of genes involved in muscle branched-chain amino acids metabolism
María Isabel Hernández-Alvarez, Angels Díaz-Ramos, María Berdasco, Jeff Cobb, Evarist Planet, Diane Cooper, Agnieszka Pazderska, Krzystof Wanic, Declan O’Hanlon, Antonio Gomez, Laura R. de la Ballina, Manel Esteller, Manuel Palacin, Donal J. O’Gorman, Joh
Scientific Reports.2017;[Epub] CrossRef - Therapeutic Targeting of the Pyruvate Dehydrogenase Complex/Pyruvate Dehydrogenase Kinase (PDC/PDK) Axis in Cancer
Peter W Stacpoole
JNCI: Journal of the National Cancer Institute.2017;[Epub] CrossRef - Hyperpalatable Diet and Physical Exercise Modulate the Expression of the Glial Monocarboxylate Transporters MCT1 and 4
Luis V. Portela, Andressa W. Brochier, Clarissa B. Haas, Afonso Kopczynski de Carvalho, Jussania A. Gnoato, Eduardo R. Zimmer, Eduardo Kalinine, Luc Pellerin, Alexandre P. Muller
Molecular Neurobiology.2017; 54(8): 5807. CrossRef - Mitochondrial targeting by dichloroacetate improves outcome following hemorrhagic shock
Kumar Subramani, Sumin Lu, Marie Warren, Xiaogang Chu, Haroldo A. Toque, R. William Caldwell, Michael P. Diamond, Raghavan Raju
Scientific Reports.2017;[Epub] CrossRef - The effect of age and unilateral leg immobilization for 2 weeks on substrate utilization during moderate‐intensity exercise in human skeletal muscle
A. Vigelsø, M. Gram, R. Dybboe, A. B. Kuhlman, C. Prats, P. L. Greenhaff, D. Constantin‐Teodosiu, J. B. Birk, J. F. P. Wojtaszewski, F. Dela, J. W. Helge
The Journal of Physiology.2016; 594(8): 2339. CrossRef - Global Kinetic Analysis of Mammalian E3 Reveals pH-dependent NAD+/NADH Regulation, Physiological Kinetic Reversibility, and Catalytic Optimum
Michael A. Moxley, Daniel A. Beard, Jason N. Bazil
Journal of Biological Chemistry.2016; 291(6): 2712. CrossRef - Pyruvate dehydrogenase kinase regulates hepatitis C virus replication
Gwon-Soo Jung, Jae-Han Jeon, Yeon-Kyung Choi, Se Young Jang, Soo Young Park, Sung-Woo Kim, Jun-Kyu Byun, Mi-Kyung Kim, Sungwoo Lee, Eui-Cheol Shin, In-Kyu Lee, Yu Na Kang, Keun-Gyu Park
Scientific Reports.2016;[Epub] CrossRef - Altered heart proteome in fructose-fed Fisher 344 rats exposed to bisphenol A
S.A. Ljunggren, M. Iggland, M. Rönn, L. Lind, P.M. Lind, H. Karlsson
Toxicology.2016; 347-349: 6. CrossRef - Unacylated ghrelin restores insulin and autophagic signaling in skeletal muscle of diabetic mice
Bjorn T. Tam, Xiao M. Pei, Benjamin Y. Yung, Shea P. Yip, Lawrence W. Chan, Cesar S. Wong, Parco M. Siu
Pflügers Archiv - European Journal of Physiology.2015; 467(12): 2555. CrossRef - Sex-related differences in the effects of high-fat diets on DHEA-treated rats
Ana Lúcia Cecconello, Marcia Trapp, Ana Lúcia Hoefel, Cláudia Vieira Marques, Bruno Dutra Arbo, Gabriela Osterkamp, Luiz Carlos Rios Kucharski, Maria Flávia Marques Ribeiro
Endocrine.2015; 48(3): 985. CrossRef - Pharmacological Blockade of Cannabinoid CB1 Receptors in Diet-Induced Obesity Regulates Mitochondrial Dihydrolipoamide Dehydrogenase in Muscle
Sergio Arrabal, Miguel Angel Lucena, Miren Josune Canduela, Almudena Ramos-Uriarte, Patricia Rivera, Antonia Serrano, Francisco Javier Pavón, Juan Decara, Antonio Vargas, Elena Baixeras, Mercedes Martín-Rufián, Javier Márquez, Pedro Fernández-Llébrez, Bau
PLOS ONE.2015; 10(12): e0145244. CrossRef - A glance at … exercise and glucose uptake
Michael J. Glade, Kyl Smith
Nutrition.2015; 31(6): 893. CrossRef - Perpetual muscle PDH activation in PDH kinase knockout mice protects against high-fat feeding–induced muscle insulin resistance
Dumitru Constantin-Teodosiu, Francis B. Stephens, Paul L. Greenhaff
Proceedings of the National Academy of Sciences.2015;[Epub] CrossRef - Statin-Induced Increases in Atrophy Gene Expression Occur Independently of Changes in PGC1α Protein and Mitochondrial Content
Craig A. Goodman, Derk Pol, Evelyn Zacharewicz, Robert S. Lee-Young, Rod J. Snow, Aaron P. Russell, Glenn K. McConell, Ashok Kumar
PLOS ONE.2015; 10(5): e0128398. CrossRef -
13C metabolic flux analysis shows that resistin impairs the metabolic response to insulin in L6E9 myotubes
Shirley Guzmán, Silvia Marin, Anibal Miranda, Vitaly A Selivanov, Josep J Centelles, Romain Harmancey, Fatima Smih, Annie Turkieh, Yves Durocher, Antonio Zorzano, Philippe Rouet, Marta Cascante
BMC Systems Biology.2014;[Epub] CrossRef - Translational Research: From Biological Discovery to Public Benefit (or Not)
Michael R. Emmert-Buck
Advances in Biology.2014; 2014: 1. CrossRef - Diabetic Worker with History of Falls: A Case Study
Ann R. Lurati
Workplace Health & Safety.2014; 62(5): 175. CrossRef - Diabetic Worker With History of Falls: A Case Study
Ann R. Lurati
Workplace Health & Safety.2014; 62(5): 175. CrossRef - Mitochondrial Pyruvate Carrier 2 Hypomorphism in Mice Leads to Defects in Glucose-Stimulated Insulin Secretion
Patrick A. Vigueira, Kyle S. McCommis, George G. Schweitzer, Maria S. Remedi, Kari T. Chambers, Xiaorong Fu, William G. McDonald, Serena L. Cole, Jerry R. Colca, Rolf F. Kletzien, Shawn C. Burgess, Brian N. Finck
Cell Reports.2014; 7(6): 2042. CrossRef
 PubReader
PubReader Cite
Cite