- Current
- Browse
- Collections
-
For contributors
- For Authors
- Instructions to authors
- Article processing charge
- e-submission
- For Reviewers
- Instructions for reviewers
- How to become a reviewer
- Best reviewers
- For Readers
- Readership
- Subscription
- Permission guidelines
- About
- Editorial policy
Articles
- Page Path
- HOME > Diabetes Metab J > Volume 38(5); 2014 > Article
-
ReviewPathophysiology Molecular Links between Caloric Restriction and Sir2/SIRT1 Activation
- Yu Wang
-
Diabetes & Metabolism Journal 2014;38(5):321-329.
DOI: https://doi.org/10.4093/dmj.2014.38.5.321
Published online: October 17, 2014
Department of Pharmacology and Pharmacy, The University of Hong Kong, Hong Kong, China.
- Corresponding author: Yu Wang. Department of Pharmacology and Pharmacy, The University of Hong Kong, Level 2, Laboratory Block, LKS Faculty of Medicine Complex, 21 Sassoon Road, Pokfulam, Hong Kong, China. yuwanghk@hku.hk
Copyright © 2014 Korean Diabetes Association
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
ABSTRACT
- Ageing is the most significant risk factor for a range of prevalent diseases, including cancer, cardiovascular disease, and diabetes. Accordingly, interventions are needed for delaying or preventing disorders associated with the ageing process, i.e., promotion of healthy ageing. Calorie restriction is the only nongenetic and the most robust approach to slow the process of ageing in evolutionarily divergent species, ranging from yeasts, worms, and flies to mammals. Although it has been known for more than 80 years that calorie restriction increases lifespan, a mechanistic understanding of this phenomenon remains elusive. Yeast silent information regulator 2 (Sir2), the founding member of the sirtuin family of protein deacetylases, and its mammalian homologue Sir2-like protein 1 (SIRT1), have been suggested to promote survival and longevity of organisms. SIRT1 exerts protective effects against a number of age-associated disorders. Caloric restriction increases both Sir2 and SIRT1 activity. This review focuses on the mechanistic insights between caloric restriction and Sir2/SIRT1 activation. A number of molecular links, including nicotinamide adenine dinucleotide, nicotinamide, biotin, and related metabolites, are suggested to be the most important conduits mediating caloric restriction-induced Sir2/SIRT1 activation and lifespan extension.
- Life expectancy has remarkably increased during the past century, due mainly to medical and pharmaceutical advances, which help to reduce life-threatening and ageing-associated diseases. For example, the discovery of antimicrobial therapy and vaccines resulted in a huge drop of infectious diseases and a major gain in life expectancy in 1900s. The modern drug development and better treatment contribute to over 50% decrease in death rates for heart disease and stroke since 1972. In 21st century, epidemics of chronic diseases, such as diabetes, obesity, cardiovascular disease, and cancer, are the targets of antiageing therapies. Increased lifespan and ageing-related problems have been brought to the forefront not only because of the financial burden to the health care and government pension system, but also due to the impacts on our society, family, and industry. In fact, many countries are facing the challenges to accommodate older workforce and to extend work lives. Innovations and new conceptions in medicine are inspiring hopes to break and further extend the biological limit of life expectancy, beyond the success achieved for lifespan extension since 1900s, especially to counteract the wide array of contemporary problems in the current century. In order for modern pharmaceutics to break the biological ageing barrier and to reach the ultimate goal of medicine, immortality, a thorough understanding of the biological basis of ageing and lifespan extension is critically urgent and important. Here, the biological pathways mediating caloric restriction-induced lifespan extension will be reviewed and discussed.
INTRODUCTION
- The lower intake of calories, the longer lifespan can be achieved. Caloric restriction without malnutrition is a nongenetic intervention that consistently promotes the extension of maximum lifespan in model organisms including yeast, worms, flies, mice, and nonhuman primates [1,2,3]. The effect can be robustly achieved by restricting up to half of the typical calorie intake in these model organisms, when malnutrition is avoided. The most striking benefit of caloric restriction is to prevent the development of a broad spectrum of ageing-associated pathological changes, such as tumorigenesis, immunosenescence and cardiometabolic disorders. In humans, long life expectancy of Okinawans is attributed to a low caloric intake and negative energy balance at younger ages, a life-long low body mass index, and a low risk of mortality from age-related diseases [4]. Optimal nutrient composition and feeding regimen of the lifespan-extending diets are not yet established. There are also debates on whether the ingested energy, when expressed in per gram body weight of the organisms, is restricted, increased or remain similar during dietary restriction [5]. In fact, the antiageing effect of caloric restriction may be achieved through restriction of certain types of amino acids, carbohydrates, lipids, or vitamins. Thus, the term 'dietary restriction' is increasingly utilized when describing limited food intake in relation to the extension of healthspan and lifespan.
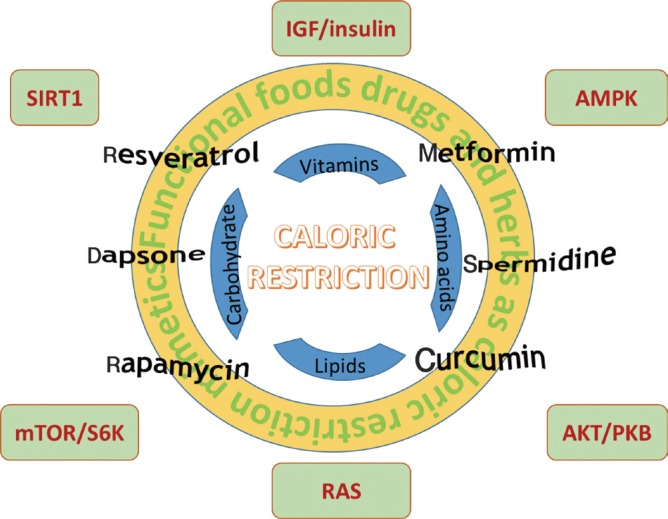
- The biological basis of caloric restriction remains poorly understood. The involvement of a single gene and pathway has been investigated in non-mammalian systems [3]. For example, removal of ethanol and/or acetic acid extends the chronological longevity (the survival of a population of nondividing cells) of the model organism yeast, whereas their replicative lifespan (the number of daughter cells generated by a single mother cell) is more sensitive to glucose restriction [6]. Down-regulation of Sch9, a serine-threonine kinase that shares high sequence identity with the mammalian Akt/protein kinase B (PKB) and ribosomal protein S6 kinase (S6K), extends the chronological lifespan by up to 2-fold [7]. Reduction of the TOR complex 1 activity leads to an extension of yeast replicative lifespan that cannot be further promoted by caloric restriction [8]. In the fruit fly Drosophila, reduction of amino acid consumption, but not sugar intake, extends life span substantially with essential amino acids mediating most of the responses [9]. In mammals, although different nutrient contents are sensed by distinctive pathways; however, it is unlikely that one single pathway is responsible for the effect of caloric restriction. Restricted dietary intake triggers the inactivation or activation of a number of nutrient sensing pathways, including insulin-like growth factor (IGF)/insulin, mammalian target of rapamycin/S6K, and silent information regulator 2 (Sir2)-like protein 1 (SIRT1) signaling pathways. These pathways are also involved in the antiageing effects of a number of chemical compounds and drugs (Fig. 1).
- In rodents, dietary restriction significantly delays the occurrence of many chronic diseases and increases life span by up to 60% [10]. Attenuated IGF-1 signaling mediates some of the antiageing effects. Excess nutrient intake activates the proageing IGF signaling pathway. Mice that are under restricted dietary intake display hypoinsulinemia, enhanced sensitivity to insulin and reduced glucose levels. In humans, dietary restriction provides similar metabolic and cardiovascular benefits as in rodents, but without reducing IGF-1 levels, unless protein intake is also reduced [11], suggesting that restriction of protein intake provides additional antiageing benefit. Older Okinawans consumed a diet with restricted calorie (10% to 15%) and low saturated fat content, but rich in functional foods (e.g., herbs or spices) that may mimic the biological effects of caloric restriction. However, the caloric restriction mimetics and the related nutrient sensing pathways remain to be characterized. In addition to longevity, caloric restriction leads to additional phenotypes, such as increased resistance to oxidative stress, enhanced repairing of DNA and protein damages, improved glucose homeostasis and insulin sensitivity, lowered serum glucose and cholesterol levels, decreased oxygen consumption and body temperature, all of which contribute to delayed onset of age-related diseases [10,12,13]. In the following part of this review, a more specific molecular mechanism involving caloric restriction-evoked activation of SIRT1 will be discussed.
CALORIC RESTRICTION AND LIFESPAN EXTENSION
- Sirtuins are a family of nicotinamide adenine dinucleotide (NAD)-dependent protein deacetylases [14]. Sir2, the first gene discovered in this family, was originally shown to regulate transcriptional silencing at cell-mating loci, telomeres, and ribosomal DNA (rDNA) in yeast, through deacetylation of the epsilon-amino groups of lysines in the amino-terminal domains of histones [15]. Sir2 cleaves the glycosidic bond between nicotinamide and adenosine diphosphate (ADP)-ribose in NAD and this reaction requires the presence of acetylated lysine [16,17]. Thus, one molecule of NAD and one molecule of acetyl-lysine are catalyzed to one molecule each of deacetylated lysine, nicotinamide and O-acetyl-ADP-ribose. There are seven mammalian sirtuins, termed SIRT1-7, which share the sequence homology of catalytic domain with Sir2 [18]. SIRT1 is the mammalian ortholog most highly related to Sir2. However, unlike the intra-nuclear localization of yeast Sir2, SIRT1 is not tightly bound to chromatin but shuttles between cytoplasm and nucleus [19]. Thus, in addition to histones, SIRT1 interacts with and mediates the deacetylation of a wide range of signaling molecules, including transcription factors, enzymes and tumor suppressors. The dynamic functions of SIRT1 are largely attributed to the additional domains at its NH2- and COOH-terminus, which allow the regulation of this protein by various posttranslational modifications and protein-protein interactions [14,20].
- During the past decade, sirtuins have attracted major attention due to their potentials of expanding life span in lower organisms and protecting against age-associated disorders in mammals. In yeast, integration of extra copies of Sir2 extends lifespan by up to 30% and deletion of this gene shortens life span by about 50% [21]. Calorie restriction by limiting glucose availability in the growth medium of the budding yeast Saccharomyces cerevisiae leads to the activation of Sir2 and the extension of replicative lifespan [22]. Sir2 mediates caloric restriction-induced lifespan extension, which requires NAD synthesis. Under conditions of reduced glucose, the metabolism of S. cerevisiae shifts from fermentation to respiration, resulting in elevated NAD or decreased NADH levels. In respiratory deficient yeast cells, on the other hand, caloric restriction could not increase Sir2 activity, but extends the replicative lifespan independent of Sir2 [23]. Moreover, the chronological lifespan of S. cerevisiae is not affected by Sir2 deficiency [24,25]. In some long-lived yeast mutants, deletion of Sir2 enhances chronological longevity extension by caloric restriction [24]. Thus, Sir2 regulates longevity in yeast through a pathway related to caloric restriction, but a direct link between these two anti-ageing factors has not been firmly demonstrated. The localization of Sir2 to certain age-related loci, such as the rDNA repeats, promotes its lifespan-sustaining function during caloric restriction [26]. On the other hand, Sir2 function is not a limiting factor for chronological ageing [27].
- The role of Sir2 in caloric restriction-induced lifespan extension has subsequently been confirmed in Caenorhabditis elegans and Drosophila melanogaster [28,29]. Unlike yeast, most cells in these fully grown and multicellular metazoan organisms are nondividing. Sir2 mediates the beneficial effects of caloric restriction via mechanisms involving metabolic control and stress responses to genotoxicity, heat shock, and oxidative damage [30,31]. Sir2 promotes genomic silencing either by repressing genomic instability or by preventing inappropriate gene expressions. In mammals, increased SIRT1 expression and function contributes to the beneficial effects of caloric restriction on delaying the onset of age-associated diseases, including cancer, atherosclerosis, and diabetes [32,33]. Mice lacking both copies of SIRT1 fail to show an increased activity and extended lifespan in response to caloric restriction, but display a shorter median lifespan than wild type mice [34,35]. Mice with elevated SIRT1 expression exhibit a beneficial phenotype resembling that of caloric restriction: they are leaner, more metabolically active, more glucose tolerant, and have reduced levels of circulating cholesterol, proinflammatory adipokines, insulin, and fasting glucose [36]. However, mice lacking one allele of SIRT1 still show identical lifespan to that observed in wild-type mice, when subjected to caloric restriction [37]. Small molecule activators of SIRT1 replicate signaling pathways triggered by calorie restriction [38]. SIRT1 plays an important role in adjusting the metabolic processes during caloric restriction, thus having been regarded as a metabolic regulator of energy homeostasis [14,39,40,41]. Importantly, SIRT1 is modulated by caloric restriction in a tissue-specific manner [42]. The activity and expression of SIRT1 in liver is reduced by caloric restriction, while those of white adipose tissue and skeletal muscle are enhanced. The systemic regulation of mammalian SIRT1 is mediated, in part, by insulin and IGF-1, two serum factors negatively involved in life-span regulation [32]. It is tentative to speculate that in tissues containing rapidly dividing cells, caloric restriction-induced SIRT1 expression shifts the balance away from cell death toward cell survival and/or regeneration, whereas in tissues containing mainly postmitotic cells, SIRT1 promotes longevity by regulating metabolic shift from using different carbon source of nutrients.
SIRTUINS AND CALORIC RESTRICTION
- When cells have high levels of calories, a substantial portion of the NAD pool is recruited into a high carbon flow of glycolysis by the enzyme glyceraldehyde-3-phosphate dehydrogenase. When calories are restricted, more carbons are oxidized in mitochondria via the electron transport chain-mediated cellular respiration, which produces NAD from NADH [43,44]. Thus, under caloric restriction, the NADH levels are significantly decreased as a result of up-regulated mitochondrial respiration [45,46]. Sir2 depletion does not affect caloric restriction-induced elevation of the intracellular NAD/NADH ratios in yeast. Because the inner membrane of mitochondria is impermeable to NADH and NAD, the malate-aspartate shuttle is used for translocating electrons produced during glycolysis for oxidative phosphorylation. This allows the hydrogen ions of NADH produced in the cytosol to reach the electron transport chain in the mitochondria. Overexpression of the malate-aspartate NADH shuttle components extends yeast replicative life span in a Sir2-dependent manner [47]. Consistently, overexpression of the mitochondrial NADH dehydrogenase specifically lowers NADH levels and extends lifespan [45]. The major modification catalyzed by Sir2/SIRT1 is deacetylation. NADH, nicotinamide adenine dinucleotide phosphate (NADP), or NADPH could not substitute NAD for this reaction. However, NAD levels do not correlate with the lifespan of yeast [46]. During calorie restriction, the NAD levels in yeast are actually decreased, indicating that Sir2 is not primarily regulated by the availability of NAD. In this regard, the deacetylase activity of Sir2 is closely linked to the decreased NADH, as the latter is a competitive inhibitor of Sir2 [45]. However, overexpressing the NADH oxidase or alternative oxidase, both of which increase NADH oxidation, could not alter the life span of the wild type yeasts [6]. These information suggest that increased respiration plays a major role in lifespan extension by caloric restriction in yeast. Sir2 acts to facilitate this process by detoxifying oxidized macromolecules, including nucleic acids, proteins and lipids. However, the activity of Sir2 and SIRT1 are not affected by physiological alterations in the NAD/NADH ratio [46].
- Unlike mitochondria, the nuclear envelope is permeable to a wide variety of small molecules [48], suggesting that cellular perturbations of NAD/NADH affect their levels in cytoplasmic as well as in nuclear compartments. NAD and NADH in nuclei play active roles in regulating gene transcription and genome stability [49]. Genotoxic stress depletes the nuclear and cytosolic pools of NAD, but not the mitochondrial pools, due largely to the extensive use of this substrate by poly (ADP-ribose) polymerases (PARPs) [50]. PARPs catalyze the polymerization of ADP-ribose units from donor NAD molecules on target proteins, resulting in the attachment of linear or branched polymers. There is a strong positive correlation between the longevity of a species and the polymer synthesis capacity of PARPs in mammalian cells [51]. PARP1 knockout mice age much faster than the wild-type control animals [52]. However, hyperactivation of PARP1 results in the depletion of NAD/adenosine triphosphate (ATP) and increases mitochondrial pore formation and cell death [53]. SIRT1 is a consumer of NAD and competes with other NAD-dependent enzymes for this common substrate. Thus, it has been proposed that inhibition of PARPs can increase NAD availability for SIRT1 to elicit the anti-ageing activity [54]. However, only certain types of tissues in PARP1 knockout mice exhibit increased NAD+ content and enhanced SIRT1 activity [55,56]. Moreover, the relationships between PARP and caloric restriction remain uncharacterized.
- Alternatively, caloric restriction may activate Sir2 by regulating the level of nicotinamide, a known inhibitor of Sir2 [57,58,59]. Crystal structures of the conserved sirtuin catalytic domains reveal that NAD and the peptide containing an acetylated lysine residue enter the active site from opposite sides of a cleft between a large Rossmann fold domain and a small Zn-binding domain [60]. During the formation of an alkylimidate intermediate between the ADP-ribose 1' position and the acetyl oxygen, nicotinamide dissociates from NAD and occupies a so-called C-pocket. If nicotinamide binds to the C-site before alkylimidate conversion, it will inhibit the deacetylation reaction. Thus, removal of nicotinamide may be as important for the activation of Sir2/SIRT1 as the production of NAD. The salvage pathway for NAD biosynthesis begins with either nicotinamide or nicotinic acid, collectively referred to as niacin or vitamin B3 [61]. Nicotinamide is first converted to nicotinamide mononucleotide (NMN) by nicotinamide phosphoribosyltransferase (NAMPT). The production of NAD+ from NMN and ATP is catalyzed by a family of nicotinamide mononucleotide adenylyltransferases (NMNATs). In lower eukaryotes, including S. cerevisiae, D. melanogaster, and C. elegans, no NAMPT activity has been found. Nicotinamide is converted to nicotinic acid, which then enters the parallel salvage pathway found in all eukaryotic species. Alternatively, nicotinamide riboside forms a precursor for NAD synthesis, connecting to the nicotinamide salvage pathway through NMN [62]. The predominant form of NMNAT in mammals, NMNAT-1, is a nuclear protein, while other forms, NMNAT-2 and NMNAT-3, are cytoplasmic and mitochondrial, respectively [63]. Overexpression of NMNAT-1 in mammalian cells does not affect total NAD levels, but regulate nuclear NAD-dependent processes [64]. Manipulation of a nuclear NAD salvage pathway delays ageing in yeast, without changing the steady-state levels of NAD [65]. Despite these information, the detailed links between caloric restriction and the NAD salvage pathway in mediating Sir2/SIRT1 activation has not been established. It is also possible that the genes involved in NAD salvage pathway act in a more general manner to promote cell survival [59,66,67].
- We have recently shown that SIRT1 is strongly inhibited by biotin, the water-soluble vitamin B7, and its metabolite biotinyl-5'-AMP [39]. Biotin occupies the binding pocket of nicotinamide, which may affect the conformational change from nonproductive to productive SIRT1 [68,69]. Biotinyl-5'-AMP competitively occupies the NAD binding site and prevents the breakdown of NAD by SIRT1. In addition, biotin may react with NAD to generate biotinyl-5'-AMP, in turn inhibiting the deacetylase activity of SIRT1. Since NAD also acts as a cofactor permitting SIRT1 to interact with protein substrates, by inhibition of NAD binding, biotinyl-5'-AMP prevents the interactions between SIRT1 and acetylated protein substrates. Adipose tissue represents a major reservoir of biotin in mammals. During ageing, biotin is progressively accumulated in adipose tissues. Chronic biotin supplementation mainly increases adipose biotin contents and abolishes adipose SIRT1-mediated beneficial effects on insulin sensitivity, lipid metabolism, and locomotor activity. In fact, caloric restriction prevents biotin accumulation in adipose tissues. Biotin and nicotinamide were originally discovered as the same class of heat-stable vitamins [70]. However, unlike nicotinamide, nutritional deficiencies of biotin are rare. The role of biotin in metabolism has been established in experimental microorganisms and animals. Biotin functions in mammals as a CO2 carrier for reactions in which a carboxyl group is transferred to one of four biotin-dependent carboxylases. Consequently, biotin participates as an important cofactor in gluconeogenesis, fatty acid synthesis, and branched-chain amino acid catabolism [71]. Based on these information, we speculate that in mammals, caloric restriction may enhance SIRT1 activity by selective depletion of biotin storage in adipose tissue, in turn preventing ageing-associated metabolic disorders and promoting lifespan extension.
MECHANISTIC INSIGHTS OF SIRT1 ACTIVATION BY CALORIC RESTRICTION
- Caloric restriction has been considered as a robust means of reducing ageing-related diseases and slowing the ageing process. Sir2 and its mammalian homologue SIRT1 are up-regulated by caloric restriction. Thus, Sir2/SIRT1 proteins sense low calories and mediate the beneficial effects of caloric restriction. However, the mechanism underlying caloric restriction-induced Sir2/SIRT1 activation remains elusive. Here, based on the available literature and our own research data, it is postulated that the induction of Sir2/SIRT1 activity by caloric restriction is an evolutionarily conserved response to decreased availability of certain nutrients, such as B vitamins.
- The initial breakthrough of identification of Sir2 as a deacetylase with weak ADP-ribosyltransferase activity came along with the identification of the Salmonella typhimurium CobB protein as a Sir2 homolog [72]. CobB compensates for the lack of CobT mutants during vitamin B12 biosynthesis and possesses nicotinate mononucleotide (NaMN)-dependent phosphoribosyltransferase activity. Thus, CobB catalyze the release of nicotinic acid from NaMN, whereas Sir2/SIRT1 removes nicotinamide from NAD. Taken together, Sir2 family of proteins play important roles in modulating the biosynthesis of B vitamins. As a feedback mechanism, increased B vitamins may negatively regulates the enzymatic activities of Sir2/SIRT1. In this regard, caloric restriction-mediated activation of Sir2/SIRT1 may at least partly relate to the nutrient availability of B vitamins, including biotin and niacin (Fig. 2).
- Calorically restricted organisms are protected from ageing-induced damages as a result of heightened defensing and repairing capacity. Various caloric restriction mimetics, including caffeine, curcumin, dapsone, metformin, rapamycin, resveratrol, and spermidine, have been developed or under development. However, none of them elicits consistent effects as caloric restriction on extending lifespan across all different organisms. The mechanisms of ageing are more complex than any single type of ageing-related diseases. It is not known which physiological changes elicited by caloric restriction in mammals are most important for longevity. The belief that many benefits of caloric restriction are due to the induction and activation of sirtuins has led to the search for promising sirtuin activators as dietary supplements to promote health and longevity. In the meantime, limiting the negative regulators of Sir2/SIRT1 by restricted diet intake may be alternative or more effective approaches.
CONCLUSIONS
-
Acknowledgements
- This work was supported by grants from Seeding Funds for Basic Research of the University of Hong Kong, Research Grant Council grants (HKU780613M, HKU779712M, HKU 781311M) and Collaborative Research Funds (HKU4/CRF/10) from Hong Kong government.
ACKNOWLEDGMENTS
- 1. Mattison JA, Roth GS, Beasley TM, Tilmont EM, Handy AM, Herbert RL, Longo DL, Allison DB, Young JE, Bryant M, Barnard D, Ward WF, Qi W, Ingram DK, de Cabo R. Impact of caloric restriction on health and survival in rhesus monkeys from the NIA study. Nature 2012;489:318-321. ArticlePubMedPDF
- 2. Colman RJ, Anderson RM, Johnson SC, Kastman EK, Kosmatka KJ, Beasley TM, Allison DB, Cruzen C, Simmons HA, Kemnitz JW, Weindruch R. Caloric restriction delays disease onset and mortality in rhesus monkeys. Science 2009;325:201-204. ArticlePubMedPMC
- 3. Fontana L, Partridge L, Longo VD. Extending healthy life span: from yeast to humans. Science 2010;328:321-326. ArticlePubMedPMC
- 4. Willcox BJ, Willcox DC, Todoriki H, Fujiyoshi A, Yano K, He Q, Curb JD, Suzuki M. Caloric restriction, the traditional Okinawan diet, and healthy aging: the diet of the world's longest-lived people and its potential impact on morbidity and life span. Ann N Y Acad Sci 2007;1114:434-455. PubMed
- 5. Yamada Y, Colman RJ, Kemnitz JW, Baum ST, Anderson RM, Weindruch R, Schoeller DA. Long-term calorie restriction decreases metabolic cost of movement and prevents decrease of physical activity during aging in rhesus monkeys. Exp Gerontol 2013;48:1226-1235. ArticlePubMed
- 6. Wei M, Fabrizio P, Madia F, Hu J, Ge H, Li LM, Longo VD. Tor1/Sch9-regulated carbon source substitution is as effective as calorie restriction in life span extension. PLoS Genet 2009;5:e1000467ArticlePubMedPMC
- 7. Fabrizio P, Pozza F, Pletcher SD, Gendron CM, Longo VD. Regulation of longevity and stress resistance by Sch9 in yeast. Science 2001;292:288-290. ArticlePubMed
- 8. Kaeberlein M, Powers RW 3rd, Steffen KK, Westman EA, Hu D, Dang N, Kerr EO, Kirkland KT, Fields S, Kennedy BK. Regulation of yeast replicative life span by TOR and Sch9 in response to nutrients. Science 2005;310:1193-1196. ArticlePubMed
- 9. Grandison RC, Piper MD, Partridge L. Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila. Nature 2009;462:1061-1064. ArticlePubMedPMCPDF
- 10. Anderson RM, Shanmuganayagam D, Weindruch R. Caloric restriction and aging: studies in mice and monkeys. Toxicol Pathol 2009;37:47-51. ArticlePubMedPDF
- 11. Fontana L, Klein S. Aging, adiposity, and calorie restriction. JAMA 2007;297:986-994. ArticlePubMed
- 12. Sohal RS, Weindruch R. Oxidative stress, caloric restriction, and aging. Science 1996;273:59-63. ArticlePubMedPMC
- 13. Park SK, Prolla TA. Lessons learned from gene expression profile studies of aging and caloric restriction. Ageing Res Rev 2005;4:55-65. ArticlePubMed
- 14. Wang Y, Xu C, Liang Y, Vanhoutte PM. SIRT1 in metabolic syndrome: where to target matters. Pharmacol Ther 2012;136:305-318. ArticlePubMed
- 15. Brachmann CB, Sherman JM, Devine SE, Cameron EE, Pillus L, Boeke JD. The SIR2 gene family, conserved from bacteria to humans, functions in silencing, cell cycle progression, and chromosome stability. Genes Dev 1995;9:2888-2902. ArticlePubMed
- 16. Landry J, Sutton A, Tafrov ST, Heller RC, Stebbins J, Pillus L, Sternglanz R. The silencing protein SIR2 and its homologs are NAD-dependent protein deacetylases. Proc Natl Acad Sci U S A 2000;97:5807-5811. ArticlePubMedPMC
- 17. Tanner KG, Landry J, Sternglanz R, Denu JM. Silent information regulator 2 family of NAD-dependent histone/protein deacetylases generates a unique product, 1-O-acetyl-ADP-ribose. Proc Natl Acad Sci U S A 2000;97:14178-14182. ArticlePubMedPMC
- 18. Haigis MC, Sinclair DA. Mammalian sirtuins: biological insights and disease relevance. Annu Rev Pathol 2010;5:253-295. ArticlePubMedPMC
- 19. Bai B, Liang Y, Xu C, Lee MY, Xu A, Wu D, Vanhoutte PM, Wang Y. Cyclin-dependent kinase 5-mediated hyperphosphorylation of sirtuin-1 contributes to the development of endothelial senescence and atherosclerosis. Circulation 2012;126:729-740. ArticlePubMed
- 20. Flick F, Luscher B. Regulation of sirtuin function by posttranslational modifications. Front Pharmacol 2012;3:29ArticlePubMedPMC
- 21. Kaeberlein M, McVey M, Guarente L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev 1999;13:2570-2580. ArticlePubMedPMC
- 22. Lin SJ, Defossez PA, Guarente L. Requirement of NAD and SIR2 for life-span extension by calorie restriction in Saccharomyces cerevisiae. Science 2000;289:2126-2128. ArticlePubMed
- 23. Kaeberlein M, Hu D, Kerr EO, Tsuchiya M, Westman EA, Dang N, Fields S, Kennedy BK. Increased life span due to calorie restriction in respiratory-deficient yeast. PLoS Genet 2005;1:e69ArticlePubMedPMC
- 24. Fabrizio P, Gattazzo C, Battistella L, Wei M, Cheng C, McGrew K, Longo VD. Sir2 blocks extreme life-span extension. Cell 2005;123:655-667. ArticlePubMed
- 25. Smith DL Jr, McClure JM, Matecic M, Smith JS. Calorie restriction extends the chronological lifespan of Saccharomyces cerevisiae independently of the Sirtuins. Aging Cell 2007;6:649-662. ArticlePubMed
- 26. Salvi JS, Chan JN, Pettigrew C, Liu TT, Wu JD, Mekhail K. Enforcement of a lifespan-sustaining distribution of Sir2 between telomeres, mating-type loci, and rDNA repeats by Rif1. Aging Cell 2013;12:67-75. ArticlePubMed
- 27. Kennedy BK, Smith ED, Kaeberlein M. The enigmatic role of Sir2 in aging. Cell 2005;123:548-550. ArticlePubMed
- 28. Rogina B, Helfand SL. Sir2 mediates longevity in the fly through a pathway related to calorie restriction. Proc Natl Acad Sci U S A 2004;101:15998-16003. ArticlePubMedPMC
- 29. Wang Y, Tissenbaum HA. Overlapping and distinct functions for a Caenorhabditis elegans SIR2 and DAF-16/FOXO. Mech Ageing Dev 2006;127:48-56. ArticlePubMed
- 30. Raynes R, Leckey BD Jr, Nguyen K, Westerheide SD. Heat shock and caloric restriction have a synergistic effect on the heat shock response in a sir2.1-dependent manner in Caenorhabditis elegans. J Biol Chem 2012;287:29045-29053. ArticlePubMedPMC
- 31. Burnett C, Valentini S, Cabreiro F, Goss M, Somogyvari M, Piper MD, Hoddinott M, Sutphin GL, Leko V, McElwee JJ, Vazquez-Manrique RP, Orfila AM, Ackerman D, Au C, Vinti G, Riesen M, Howard K, Neri C, Bedalov A, Kaeberlein M, Soti C, Partridge L, Gems D. Absence of effects of Sir2 overexpression on lifespan in C. elegans and Drosophila. Nature 2011;477:482-485. ArticlePubMedPMCPDF
- 32. Cohen HY, Miller C, Bitterman KJ, Wall NR, Hekking B, Kessler B, Howitz KT, Gorospe M, de Cabo R, Sinclair DA. Calorie restriction promotes mammalian cell survival by inducing the SIRT1 deacetylase. Science 2004;305:390-392. ArticlePubMed
- 33. Herranz D, Munoz-Martin M, Canamero M, Mulero F, Martinez-Pastor B, Fernandez-Capetillo O, Serrano M. Sirt1 improves healthy ageing and protects from metabolic syndrome-associated cancer. Nat Commun 2010;1:3ArticlePubMedPDF
- 34. Chen D, Steele AD, Lindquist S, Guarente L. Increase in activity during calorie restriction requires Sirt1. Science 2005;310:1641ArticlePubMed
- 35. Boily G, Seifert EL, Bevilacqua L, He XH, Sabourin G, Estey C, Moffat C, Crawford S, Saliba S, Jardine K, Xuan J, Evans M, Harper M-E, McBurney MW. SirT1 regulates energy metabolism and response to caloric restriction in mice. PLoS One 2008;3:e1759ArticlePubMedPMC
- 36. Bordone L, Cohen D, Robinson A, Motta MC, van Veen E, Czopik A, Steele AD, Crowe H, Marmor S, Luo J, Gu W, Guarente L. SIRT1 transgenic mice show phenotypes resembling calorie restriction. Aging Cell 2007;6:759-767. ArticlePubMed
- 37. Mercken EM, Hu J, Krzysik-Walker S, Wei M, Li Y, McBurney MW, de Cabo R, Longo VD. SIRT1 but not its increased expression is essential for lifespan extension in caloric-restricted mice. Aging Cell 2014;13:193-196. ArticlePubMed
- 38. Smith JJ, Kenney RD, Gagne DJ, Frushour BP, Ladd W, Galonek HL, Israelian K, Song J, Razvadauskaite G, Lynch AV, Carney DP, Johnson RJ, Lavu S, Iffland A, Elliott PJ, Lambert PD, Elliston KO, Jirousek MR, Milne JC, Boss O. Small molecule activators of SIRT1 replicate signaling pathways triggered by calorie restriction in vivo. BMC Syst Biol 2009;3:31ArticlePubMedPMCPDF
- 39. Xu C, Bai B, Fan P, Cai Y, Huang B, Law IK, Liu L, Xu A, Tung C, Li X, Siu M, Che CM, Vanhoutte PM, Wang Y. Selective overexpression of human SIRT1 in adipose tissue enhances energy homeostasis and prevents the deterioration of insulin sensitivity with ageing in mice. Am J Transl Res 2013;5:412-426. PubMedPMC
- 40. Guarente L. Sirtuins as potential targets for metabolic syndrome. Nature 2006;444:868-874. ArticlePubMedPDF
- 41. Pfluger PT, Herranz D, Velasco-Miguel S, Serrano M, Tschop MH. Sirt1 protects against high-fat diet-induced metabolic damage. Proc Natl Acad Sci U S A 2008;105:9793-9798. ArticlePubMedPMC
- 42. Chen D, Bruno J, Easlon E, Lin SJ, Cheng HL, Alt FW, Guarente L. Tissue-specific regulation of SIRT1 by calorie restriction. Genes Dev 2008;22:1753-1757. ArticlePubMedPMC
- 43. Guarente L. Sir2 links chromatin silencing, metabolism, and aging. Genes Dev 2000;14:1021-1026. ArticlePubMed
- 44. Bakker BM, Overkamp KM, van Maris AJ, Kotter P, Luttik MA, van Dijken JP, Pronk JT. Stoichiometry and compartmentation of NADH metabolism in Saccharomyces cerevisiae. FEMS Microbiol Rev 2001;25:15-37. ArticlePubMed
- 45. Lin SJ, Ford E, Haigis M, Liszt G, Guarente L. Calorie restriction extends yeast life span by lowering the level of NADH. Genes Dev 2004;18:12-16. ArticlePubMedPMC
- 46. Anderson RM, Latorre-Esteves M, Neves AR, Lavu S, Medvedik O, Taylor C, Howitz KT, Santos H, Sinclair DA. Yeast life-span extension by calorie restriction is independent of NAD fluctuation. Science 2003;302:2124-2126. ArticlePubMedPMC
- 47. Easlon E, Tsang F, Skinner C, Wang C, Lin SJ. The malate-aspartate NADH shuttle components are novel metabolic longevity regulators required for calorie restriction-mediated life span extension in yeast. Genes Dev 2008;22:931-944. ArticlePubMedPMC
- 48. Paine PL. Nucleocytoplasmic movement of fluorescent tracers microinjected into living salivary gland cells. J Cell Biol 1975;66:652-657. ArticlePubMedPMCPDF
- 49. Zhang Q, Piston DW, Goodman RH. Regulation of corepressor function by nuclear NADH. Science 2002;295:1895-1897. ArticlePubMed
- 50. Kim MY, Zhang T, Kraus WL. Poly(ADP-ribosyl)ation by PARP-1: 'PAR-laying' NAD+ into a nuclear signal. Genes Dev 2005;19:1951-1967. ArticlePubMed
- 51. Beneke S, Scherr AL, Ponath V, Popp O, Burkle A. Enzyme characteristics of recombinant poly(ADP-ribose) polymerases-1 of rat and human origin mirror the correlation between cellular poly(ADP-ribosyl)ation capacity and species-specific life span. Mech Ageing Dev 2010;131:366-369. ArticlePubMed
- 52. Piskunova TS, Yurova MN, Ovsyannikov AI, Semenchenko AV, Zabezhinski MA, Popovich IG, Wang ZQ, Anisimov VN. Deficiency in poly(ADP-ribose) polymerase-1 (PARP-1) accelerates aging and spontaneous carcinogenesis in mice. Curr Gerontol Geriatr Res 2008;2008:754190ArticlePubMedPMCPDF
- 53. Mangerich A, Burkle A. Pleiotropic cellular functions of PARP1 in longevity and aging: genome maintenance meets inflammation. Oxid Med Cell Longev 2012;2012:321653ArticlePubMedPMCPDF
- 54. El Ramy R, Magroun N, Messadecq N, Gauthier LR, Boussin FD, Kolthur-Seetharam U, Schreiber V, McBurney MW, Sassone-Corsi P, Dantzer F. Functional interplay between Parp-1 and SirT1 in genome integrity and chromatin-based processes. Cell Mol Life Sci 2009;66:3219-3234. ArticlePubMedPDF
- 55. Bai P, Canto C, Brunyanszki A, Huber A, Szanto M, Cen Y, Yamamoto H, Houten SM, Kiss B, Oudart H, Gergely P, Menissier-de Murcia J, Schreiber V, Sauve AA, Auwerx J. PARP-2 regulates SIRT1 expression and whole-body energy expenditure. Cell metabolism 2011;13:450-460. ArticlePubMedPMC
- 56. Bai P, Canto C, Oudart H, Brunyanszki A, Cen Y, Thomas C, Yamamoto H, Huber A, Kiss B, Houtkooper RH, Schoonjans K, Schreiber V, Sauve AA, Menissier-de Murcia J, Auwerx J. PARP-1 inhibition increases mitochondrial metabolism through SIRT1 activation. Cell Metab 2011;13:461-468. ArticlePubMedPMC
- 57. Landry J, Slama JT, Sternglanz R. Role of NAD(+) in the deacetylase activity of the SIR2-like proteins. Biochem Biophys Res Commun 2000;278:685-690. ArticlePubMed
- 58. Bitterman KJ, Anderson RM, Cohen HY, Latorre-Esteves M, Sinclair DA. Inhibition of silencing and accelerated aging by nicotinamide, a putative negative regulator of yeast sir2 and human SIRT1. J Biol Chem 2002;277:45099-45107. ArticlePubMed
- 59. Anderson RM, Bitterman KJ, Wood JG, Medvedik O, Sinclair DA. Nicotinamide and PNC1 govern lifespan extension by calorie restriction in Saccharomyces cerevisiae. Nature 2003;423:181-185. ArticlePubMedPMCPDF
- 60. Avalos JL, Bever KM, Wolberger C. Mechanism of sirtuin inhibition by nicotinamide: altering the NAD(+) cosubstrate specificity of a Sir2 enzyme. Mol Cell 2005;17:855-868. ArticlePubMed
- 61. Rongvaux A, Andris F, Van Gool F, Leo O. Reconstructing eukaryotic NAD metabolism. Bioessays 2003;25:683-690. ArticlePubMed
- 62. Bieganowski P, Brenner C. Discoveries of nicotinamide riboside as a nutrient and conserved NRK genes establish a Preiss-Handler independent route to NAD+ in fungi and humans. Cell 2004;117:495-502. ArticlePubMed
- 63. Berger F, Ramirez-Hernandez MH, Ziegler M. The new life of a centenarian: signalling functions of NAD(P). Trends Biochem Sci 2004;29:111-118. ArticlePubMed
- 64. Araki T, Sasaki Y, Milbrandt J. Increased nuclear NAD biosynthesis and SIRT1 activation prevent axonal degeneration. Science 2004;305:1010-1013. ArticlePubMed
- 65. Anderson RM, Bitterman KJ, Wood JG, Medvedik O, Cohen H, Lin SS, Manchester JK, Gordon JI, Sinclair DA. Manipulation of a nuclear NAD+ salvage pathway delays aging without altering steady-state NAD+ levels. J Biol Chem 2002;277:18881-18890. ArticlePubMed
- 66. Yang H, Lavu S, Sinclair DA. Nampt/PBEF/Visfatin: a regulator of mammalian health and longevity? Exp Gerontol 2006;41:718-726. ArticlePubMedPMC
- 67. Gallo CM, Smith DL Jr, Smith JS. Nicotinamide clearance by Pnc1 directly regulates Sir2-mediated silencing and longevity. Mol Cell Biol 2004;24:1301-1312. ArticlePubMedPMCPDF
- 68. Min J, Landry J, Sternglanz R, Xu RM. Crystal structure of a SIR2 homolog-NAD complex. Cell 2001;105:269-279. ArticlePubMed
- 69. Zhao K, Harshaw R, Chai X, Marmorstein R. Structural basis for nicotinamide cleavage and ADP-ribose transfer by NAD(+)-dependent Sir2 histone/protein deacetylases. Proc Natl Acad Sci U S A 2004;101:8563-8568. ArticlePubMedPMC
- 70. Lanska DJ. The discovery of niacin, biotin, and pantothenic acid. Ann Nutr Metab 2012;61:246-253. ArticlePubMedPDF
- 71. Depeint F, Bruce WR, Shangari N, Mehta R, O'Brien PJ. Mitochondrial function and toxicity: role of the B vitamin family on mitochondrial energy metabolism. Chem Biol Interact 2006;163:94-112. ArticlePubMed
- 72. Tsang AW, Escalante-Semerena JC. CobB, a new member of the SIR2 family of eucaryotic regulatory proteins, is required to compensate for the lack of nicotinate mononucleotide: 5,6-dimethylbenzimidazole phosphoribosyltransferase activity in cobT mutants during cobalamin biosynthesis in Salmonella typhimurium LT2. J Biol Chem 1998;273:31788-31794. ArticlePubMed
REFERENCES
Fig. 1In response to different dietary intake, a number of nutrient sensing pathways are activated or inactivated to modulate the ageing process. IGF, insulin-like growth factor; SIRT1, Sir2-like protein 1; AMPK, AMP-activated protein kinase; mTOR/S6K, mammalian target of rapamycin/ribosomal protein S6 kinase; ROS, reactive oxygen species; AKT/PKB, AKT/protein kinase B.
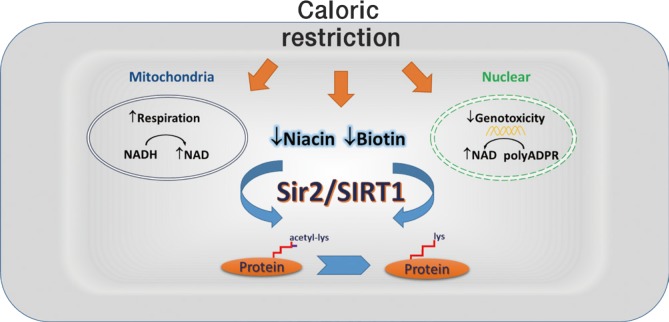
Figure & Data
References
Citations
Citations to this article as recorded by 
- Caloric Restriction Can Ameliorate Postoperative Cognitive Dysfunction by Upregulating the Expression of Sirt1, MeCP2 and BDNF in the Hippocampal CA1 Region of Aged C57BL/6 Mice
Lan Wei, Qiang Tao, Minmin Yao, Zhimeng Zhao, Shengjin Ge
Brain Sciences.2023; 13(3): 462. CrossRef - Metabolic Syndrome and Cardiac Remodeling Due to Mitochondrial Oxidative Stress Involving Gliflozins and Sirtuins
Raúl Lelio Sanz, Felipe Inserra, Sebastián García Menéndez, Luciana Mazzei, León Ferder, Walter Manucha
Current Hypertension Reports.2023; 25(6): 91. CrossRef - The Tricarboxylic Acid Cycle as a Central Regulator of the Rate of Aging: Implications for Metabolic Interventions
Jonathan M. Borkum
Advanced Biology.2023;[Epub] CrossRef - Metabolic Reprogramming by Reduced Calorie Intake or Pharmacological Caloric Restriction Mimetics for Improved Cancer Immunotherapy
Erwan Eriau, Juliette Paillet, Guido Kroemer, Jonathan G. Pol
Cancers.2021; 13(6): 1260. CrossRef - SGLT2 Inhibitors as Calorie Restriction Mimetics: Insights on Longevity Pathways and Age-Related Diseases
Caroline W S Hoong, Marvin W J Chua
Endocrinology.2021;[Epub] CrossRef - A comprehensive insight into the potential effects of resveratrol supplementation on SIRT-1: A systematic review
Marziyeh Najafi, Omid Nikpayam, Omid Mohammad Tavakoli-Rouzbehani, Shahab Papi, Atefeh Amrollahi bioky, Elham sadat Ahmadiani, Golbon Sohrab
Diabetes & Metabolic Syndrome: Clinical Research & Reviews.2021; 15(5): 102224. CrossRef - Antiaging and Antioxidant Bioactivities of Asteraceae Plant Fractions on the Cellular Functions of the Yeast Schizosaccharomyces pombe
Rika Indri Astuti, Muhammad Eka Prastya, Irmanida Batubara, Eka Budiarti, Aulia Ilmiyawati, Mounir Tilaoui
Advances in Pharmacological and Pharmaceutical Sciences.2021; 2021: 1. CrossRef - Principles of brain aging: Status and challenges of modeling human molecular changes in mice
Verena Kluever, Eugenio F. Fornasiero
Ageing Research Reviews.2021; 72: 101465. CrossRef - Role of NRF2 and Sirtuin activators in COVID-19
Hasnat Khan, Shivangi Patel, Anuradha Majumdar
Clinical Immunology.2021; 233: 108879. CrossRef - Role of acetylation in nonalcoholic fatty liver disease: a focus on SIRT1 and SIRT3
Fatiha Nassir
Exploration of Medicine.2020; 1(4): 248. CrossRef - NAD metabolism in aging and cancer
John WR Kincaid, Nathan A Berger
Experimental Biology and Medicine.2020; 245(17): 1594. CrossRef - Cyclin‐dependent kinase Pho85p and its cyclins are involved in replicative lifespan through multiple pathways in yeast
Toshio Nakajima, Tsubasa Maruhashi, Takaaki Morimatsu, Yukio Mukai
FEBS Letters.2020; 594(7): 1166. CrossRef - SIRT1 Allele Frequencies in Depressed Patients of European Descent in Russia
Lyubomir I. Aftanas, Maksim S. Anisimenko, Darya A. Berdyugina, Aleksandr Yu. Garanin, Vladimir N. Maximov, Mikhail I. Voevoda, Natalya M. Vyalova, Nikolay A. Bokhan, Svetlana A. Ivanova, Konstantin V. Danilenko, Sergei P. Kovalenko
Frontiers in Genetics.2019;[Epub] CrossRef - Resveratrol attenuates myocardial hypoxia/reoxygenation-induced cell apoptosis through DJ-1-mediated SIRT1-p53 pathway
Rui-Yuan Xu, Xing-Wang Xu, Yi-Zhang Deng, Zhao-Xia Ma, Xiao-Ran Li, Le Zhao, Le-Jia Qiu, Hao-Yue Liu, He-Ping Chen
Biochemical and Biophysical Research Communications.2019; 514(2): 401. CrossRef - The pathobiology of polycystic kidney disease from a metabolic viewpoint
Luis Fernando Menezes, Gregory G. Germino
Nature Reviews Nephrology.2019; 15(12): 735. CrossRef - Six Weeks of Calorie Restriction Improves Body Composition and Lipid Profile in Obese and Overweight Former Athletes
Joanna Hołowko, Małgorzata Magdalena Michalczyk, Adam Zając, Maja Czerwińska-Rogowska, Karina Ryterska, Marcin Banaszczak, Karolina Jakubczyk, Ewa Stachowska
Nutrients.2019; 11(7): 1461. CrossRef - Sirtuins in gamete biology and reproductive physiology: emerging roles and therapeutic potential in female and male infertility
Carla Tatone, Giovanna Di Emidio, Arcangelo Barbonetti, Gaspare Carta, Alberto M Luciano, Stefano Falone, Fernanda Amicarelli
Human Reproduction Update.2018; 24(3): 267. CrossRef - Is there a role of H2S in mediating health span benefits of caloric restriction?
Li Theng Ng, Jan Gruber, Philip Keith Moore
Biochemical Pharmacology.2018; 149: 91. CrossRef - Sirtuins and Accelerated Aging in Scleroderma
Anne E. Wyman, Sergei P. Atamas
Current Rheumatology Reports.2018;[Epub] CrossRef - Comparison of Calorie-Restricted Diet and Resveratrol Supplementation on Anthropometric Indices, Metabolic Parameters, and Serum Sirtuin-1 Levels in Patients With Nonalcoholic Fatty Liver Disease: A Randomized Controlled Clinical Trial
Somayyeh Asghari, Mohammad Asghari-Jafarabadi, Mohammad-Hossein Somi, Seyed-Mostafa Ghavami, Maryam Rafraf
Journal of the American College of Nutrition.2018; 37(3): 223. CrossRef - Amino Acid Restriction Triggers Angiogenesis via GCN2/ATF4 Regulation of VEGF and H2S Production
Alban Longchamp, Teodelinda Mirabella, Alessandro Arduini, Michael R. MacArthur, Abhirup Das, J. Humberto Treviño-Villarreal, Christopher Hine, Issam Ben-Sahra, Nelson H. Knudsen, Lear E. Brace, Justin Reynolds, Pedro Mejia, Ming Tao, Gaurav Sharma, Rui W
Cell.2018; 173(1): 117. CrossRef - Recent insights into the cellular and molecular determinants of aging
Linhao Ruan, Xi Zhang, Rong Li
Journal of Cell Science.2018;[Epub] CrossRef - SIRT1 Activity in Peripheral Blood Mononuclear Cells Correlates with Altered Lung Function in Patients with Chronic Obstructive Pulmonary Disease
Valeria Conti, Graziamaria Corbi, Valentina Manzo, Paola Malangone, Carolina Vitale, Angelantonio Maglio, Roberta Cotugno, Damiano Capaccio, Luigi Marino, Carmine Selleri, Cristiana Stellato, Amelia Filippelli, Alessandro Vatrella
Oxidative Medicine and Cellular Longevity.2018; 2018: 1. CrossRef - Sirt1: Role Under the Condition of Ischemia/Hypoxia
Xiaofei Meng, Jin Tan, Mengmeng Li, Shuling Song, Yuyang Miao, Qiang Zhang
Cellular and Molecular Neurobiology.2017; 37(1): 17. CrossRef - Salidroside protects PC12 cells from Aβ1–40-induced cytotoxicity by regulating the nicotinamide phosphoribosyltransferase signaling pathway
Xujiao Huang, Sanli Xing, Chuan Chen, Zhihua Yu, Jiulin Chen
Molecular Medicine Reports.2017; 16(3): 2700. CrossRef - Combined Salt and Caloric Restrictions: Potential Adverse Outcomes
Tsuyoshi Homma, Mika Homma, Yuefei Huang, Korapat Mayurasakorn, Nurul Mahamad Rodi, Anis Amalina Abdul Hamid, Shelley Hurwitz, Tham Yao, Gail K. Adler, Luminita H. Pojoga, Gordon H. Williams, Jose R. Romero
Journal of the American Heart Association.2017;[Epub] CrossRef - Hyperglycemic Stress and Carbon Stress in Diabetic Glucotoxicity
Xiaoting Luo, Jinzi Wu, Siqun Jing, Liang-Jun Yan
Aging and disease.2016; 7(1): 90. CrossRef - Endothelial SIRT1 prevents adverse arterial remodeling by facilitating HERC2-mediated degradation of acetylated LKB1
Bo Bai, Andy W.C. Man, Kangmin Yang, Yumeng Guo, Cheng Xu, Hung-Fat Tse, Weiping Han, Maria Bloksgaard, Jo G.R. De Mey, Paul M. Vanhoutte, Aimin Xu, Yu Wang
Oncotarget.2016; 7(26): 39065. CrossRef - Circadian clocks govern calorie restriction—mediated life span extension through BMAL1‐ and IGF‐1‐dependent mechanisms
Sonal A. Patel, Amol Chaudhari, Richa Gupta, Nikkhil Velingkaar, Roman V. Kondratov
The FASEB Journal.2016; 30(4): 1634. CrossRef - Nutrigenomics at the Interface of Aging, Lifespan, and Cancer Prevention
Gabriela Riscuta
The Journal of Nutrition.2016; 146(10): 1931. CrossRef - Haploinsufficiency of the Myc regulator Mtbp extends survival and delays tumor development in aging mice
Brian C. Grieb, Kelli Boyd, Ramkrishna Mitra, Christine M. Eischen
Aging.2016; 8(10): 2590. CrossRef - Protein Modifications as Manifestations of Hyperglycemic Glucotoxicity in Diabetes and Its Complications
Hong Zheng, Jinzi Wu, Zhen Jin, Liang-Jun Yan
Biochemistry Insights.2016; 9: BCI.S36141. CrossRef - Nutritional limitation in early postnatal life and its effect on aging and longevity in rodents
Kallie Davis, Douja Chamseddine, James M. Harper
Experimental Gerontology.2016; 86: 84. CrossRef - 1,4-Dihydropyridines Active on the SIRT1/AMPK Pathway Ameliorate Skin Repair and Mitochondrial Function and Exhibit Inhibition of Proliferation in Cancer Cells
Sergio Valente, Paolo Mellini, Francesco Spallotta, Vincenzo Carafa, Angela Nebbioso, Lucia Polletta, Ilaria Carnevale, Serena Saladini, Daniela Trisciuoglio, Chiara Gabellini, Maria Tardugno, Clemens Zwergel, Chiara Cencioni, Sandra Atlante, Sébastien Mo
Journal of Medicinal Chemistry.2016; 59(4): 1471. CrossRef - Involvement of Daphnia pulicaria Sir2 in regulating stress response and lifespan
Charles A. Schumpert, Craig Anderson, Jeffry L. Dudycha, Rekha C. Patel
Aging.2016; 8(2): 402. CrossRef - Metformin Restores Parkin-Mediated Mitophagy, Suppressed by Cytosolic p53
Young Song, Woo Lee, Yong-ho Lee, Eun Kang, Bong-Soo Cha, Byung-Wan Lee
International Journal of Molecular Sciences.2016; 17(1): 122. CrossRef - Are sirtuins markers of ovarian aging?
Jinjin Zhang, Li Fang, Zhiyong Lu, Jiaqiang Xiong, Meng Wu, Liangyan Shi, Aiyue Luo, Shixuan Wang
Gene.2016; 575(2): 680. CrossRef - Coadjuvants in the Diabetic Complications: Nutraceuticals and Drugs with Pleiotropic Effects
Thiago Pereira, Fabio Pimenta, Marcella Porto, Marcelo Baldo, Bianca Campagnaro, Agata Gava, Silvana Meyrelles, Elisardo Vasquez
International Journal of Molecular Sciences.2016; 17(8): 1273. CrossRef - Extended longevity and survivorship during amino-acid starvation in a DrosophilaSir2mutant heterozygote
Jennifer D. Slade, Brian E. Staveley, A. Civetta
Genome.2016; 59(5): 311. CrossRef - Resveratrol inhibits doxorubicin-induced cardiotoxicity via sirtuin 1 activation in H9c2 cardiomyocytes
Mi-Hua Liu, Jian Shan, Jian Li, Yuan Zhang, Xiao-Long Lin
Experimental and Therapeutic Medicine.2016; 12(2): 1113. CrossRef - Effect of caloric restriction on the SIRT1/mTOR signaling pathways in senile mice
Lina Ma, Wen Dong, Rong Wang, Yun Li, Baolei Xu, Jingshuang Zhang, Zhiwei Zhao, Yulan Wang
Brain Research Bulletin.2015; 116: 67. CrossRef - Metformin prevents aggressive ovarian cancer growth driven by high-energy diet: similarity with calorie restriction
Zaid Al-Wahab, Ismail Mert, Calvin Tebbe, Jasdeep Chhina, Miriana Hijaz, Robert T. Morris, Rouba Ali-Fehmi, Shailendra Giri, Adnan R. Munkarah, Ramandeep Rattan
Oncotarget.2015; 6(13): 10908. CrossRef - Restriction on an Energy-Dense Diet Improves Markers of Metabolic Health and Cellular Aging in Mice Through Decreasing Hepatic mTOR Activity
Anke Schloesser, Graeme Campbell, Claus-Christian Glüer, Gerald Rimbach, Patricia Huebbe
Rejuvenation Research.2015; 18(1): 30. CrossRef - Calorie Restriction Prevents Metabolic Aging Caused by Abnormal SIRT1 Function in Adipose Tissues
Cheng Xu, Yu Cai, Pengcheng Fan, Bo Bai, Jie Chen, Han-Bing Deng, Chi-Ming Che, Aimin Xu, Paul M. Vanhoutte, Yu Wang
Diabetes.2015; 64(5): 1576. CrossRef
 PubReader
PubReader Cite
Cite