- Current
- Browse
- Collections
-
For contributors
- For Authors
- Instructions to authors
- Article processing charge
- e-submission
- For Reviewers
- Instructions for reviewers
- How to become a reviewer
- Best reviewers
- For Readers
- Readership
- Subscription
- Permission guidelines
- About
- Editorial policy
Articles
- Page Path
- HOME > Diabetes Metab J > Volume 37(3); 2013 > Article
-
ReviewObesity and Metabolic Syndrome Chemokine Systems Link Obesity to Insulin Resistance
- Tsuguhito Ota
-
Diabetes & Metabolism Journal 2013;37(3):165-172.
DOI: https://doi.org/10.4093/dmj.2013.37.3.165
Published online: June 14, 2013
Department of Cell Metabolism and Nutrition, Brain/Liver Interface Medicine Research Center, Kanazawa University School of Medicine, Kanazawa, Japan.
- Corresponding author: Tsuguhito Ota. Department of Cell Metabolism and Nutrition, Brain/Liver Interface Medicine Research Center, Kanazawa University School of Medicine, 13-1 Takara-machi, Kanazawa, Ishikawa 920-8641, Japan. tota@staff.kanazawa-u.ac.jp
Copyright © 2013 Korean Diabetes Association
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
- ABSTRACT
- INTRODUCTION
- CLASSIFICATION OF CHEMOKINES AND CHEMOKINE RECEPTORS
- INFLAMMATORY CHEMOKINES AND HOMEOSTATIC CHEMOKINES
- CHEMOKINE SYSTEMS AND DISEASE
- PIVOTAL ROLE OF MCP-1-CCR2 IN OBESITY-INDUCED INSULIN RESISTANCE
- CCR5 LINKS OBESITY TO INSULIN RESISTANCE BY REGULATING BOTH MACROPHAGE RECRUITMENT AND M1/M2 STATUS
- CCR5 AS A NOVEL PLAYER IN THE ADIPOSE TISSUE INFLAMMATION AND INSULIN RESISTANCE
- CONCLUSIONS
- ACKNOWLEDGMENTS
- NOTES
- REFERENCES
ABSTRACT
- Obesity is a state of chronic low-grade systemic inflammation. This chronic inflammation is deeply involved in insulin resistance, which is the underlying condition of type 2 diabetes and metabolic syndrome. A significant advance in our understanding of obesity-associated inflammation and insulin resistance has been recognition of the critical role of adipose tissue macrophages (ATMs). Chemokines are small proteins that direct the trafficking of immune cells to sites of inflammation. In addition, chemokines activate the production and secretion of inflammatory cytokines through specific G protein-coupled receptors. ATM accumulation through C-C motif chemokine receptor 2 and its ligand monocyte chemoattractant protein-1 is considered pivotal in the development of insulin resistance. However, chemokine systems appear to exhibit a high degree of functional redundancy. Currently, more than 50 chemokines and 18 chemokine receptors exhibiting various physiological and pathological properties have been discovered. Therefore, additional, unidentified chemokine/chemokine receptor pathways that may play significant roles in ATM recruitment and insulin sensitivity remain to be fully identified. This review focuses on some of the latest findings on chemokine systems linking obesity to inflammation and subsequent development of insulin resistance.
- Excess adiposity increases the risk of developing a variety of pathological conditions, including type 2 diabetes, cardiovascular disease, steatohepatitis, and several types of cancer [1-4]. In the past two decades, advances in obesity research have led to the recognition that adipose tissue is an endocrine organ that secretes hormones or cytokines termed adipokines [5,6]. Adipokines play crucial roles in the regulation of appetite and satiety control, energy expenditure, insulin sensitivity and insulin secretion, endothelial function, and blood pressure [3,6]. The adipokines leptin and adiponectin, which are primarily secreted by adipocytes, play a major role in the pathogenesis of obesity and its comorbidities, including type 2 diabetes and cardiovascular disease [7,8]. On the other hand, obesity involves a state of chronic low-grade systemic inflammation. Recently, areas of active investigation focus on the molecular bases of metabolic inflammation and potential pathogenic roles in diabetes and cardiovascular disease [2,9,10]. This inflammation develops in response to an excess of nutrient flux into metabolic tissues including adipose tissue, liver, and skeletal muscle and is currently recognized as an important link between obesity and insulin resistance. Obesity-associated systemic inflammation is characterized by increased circulating concentrations of proinflammatory cytokines and chemokines, and activation of several kinases that regulate inflammation, including c-Jun NH2 terminal kinase, IκB-kinase β/nuclear factor κB and mammalian target of rapamycin/S6 kinase that interfere with insulin action in adipocytes and hepatocytes.
- Increasing evidence supports that obesity-induced inflammation is mediated primarily by immune cells such as the macrophages and T lymphocytes in metabolic tissues. In particular, a significant advance in our understanding of obesity-associated inflammation and insulin resistance has been recognition of the critical role of adipose tissue macrophages (ATMs). ATMs are a prominent source of proinflammatory cytokines, such as tumor necrosis factor (TNF)-α and interleukin (IL)-6, that can block insulin action in adipocytes via autocrine/paracrine signaling and cause systemic insulin resistance via endocrine signaling, providing a potential link between inflammation and insulin resistance [10-12]. In both humans and rodents, ATMs accumulate in adipose tissue with increasing body weight and their content correlates positively with insulin resistance [13-15]. Importantly, tissue macrophages are phenotypically heterogeneous and have been characterized according to their activation/polarization state as M1 (or "classically activated" proinflammatory macrophages) or M2 (or "alternatively activated" noninflammatory macrophages [16-18]). M2 ATMs predominate in lean mice, whereas obesity induces the accumulation of M1 ATMs with high expression of TNF-α, IL-6 and inducible nitric oxide synthase [17], leading to a proinflammatory environment in white adipose tissue (WAT). Thus, both recruitment and proinflammatory activation of ATMs is required for the development of insulin resistance in obese mice.
- Chemokines, a family of cytokines that induce leukocyte chemotaxis, are deeply involved in the development of allergic diseases and autoimmune diseases. More than 50 chemokines which exhibit various physiological and pathological properties have been discovered to date [16,19]. Recent studies have shown that preadipocytes and adipocytes express chemokines, and have demonstrated the infiltration of bone marrow-derived macrophages into obese adipose tissue, which is involved in the development of insulin resistance. This review summarizes various roles of chemokine and chemokine receptor (chemokine system) in inflammation, and also focuses on some of the latest findings on the chemokine systems that link adipose tissue inflammation with the pathology of insulin resistance.
INTRODUCTION
- Chemokines attract leukocytes to areas of inflammation and lesions and play a key role in leukocyte activation. Chemokines were first discovered as cytokines that are chemotactic for neutrophils and monocytes. Ever since, many studies have been conducted to identify the roles of chemokines in acute, neutrophil-predominant inflammation and chronic, monocyte- and lymphocyte-predominant inflammation [20,21]. To date, more than 50 chemokines have been identified in humans (Table 1). Chemokines have four conserved cysteine residues. Based on the motif patterns involving two N-terminal cysteine residues, chemokines can be classified into the following four subfamilies: CXC, CC, C, and CX3C (where X is any amino acid residue) (Table 1) [16,19]. The CXC chemokines are mainly chemotactic for neutrophils and are known for their involvement in acute inflammation whereas most of CC chemokines act on monocytes, T cells, eosinophils, and basophils, which mediate chronic inflammation and allergy [16,19].
- All chemokines signal via seven-transmembrane G-protein-coupled receptors (or chemokine receptors). To date, 18 chemokine receptors have been identified, including 11 CC chemokine receptors, 6 CXC chemokine receptors, and one for each C (XCR1) and CX3C chemokine receptor (Table 1). Most chemokines bind to several chemokine receptors and chemokine receptors have overlapping ligand specificities [16,19]. Although some chemokines have a one-to-one specificity (specific receptors), most chemokines bind to the same receptor (shared receptors) (Table 1) [16,19]. For example, four chemokine ligands, including monocyte chemoattractant protein (MCP)-1, MCP-2, MCP-3, and MCP-4 bind to C-C motif chemokine receptor 2 (CCR2) receptor. Even when multiple ligands interact with a single receptor, diverse effects are produced by different ligands because the binding affinity and the resulting effect differ across ligands. Furthermore, as chemokines are differently expressed, distributed, and regulated in cells and tissue, they may play different roles in physiological conditions or diseases.
CLASSIFICATION OF CHEMOKINES AND CHEMOKINE RECEPTORS
- Although chemokines appear to exhibit a high degree of functional redundancy, the functions of the individual chemokines can be classified into two types: 'inducible' or 'inflammatory' and 'constitutive' or 'homeostatic' [16,19]. Inflammatory CXC and CC chemokines promptly recruit neutrophils, monocytes and eosinophils to the site of injury or inflammation, cluster on chromosomes 4 and 17, and share the same receptor [22]. Furthermore, a number of inflammatory chemokines act together as a potent inducer of cell migration (quantitative regulation). Since the late 1990s, it has been clearly demonstrated that several chemokines exhibit different properties from those of inflammatory chemokines. These homeostatic chemokines act specifically and constantly on lymphocytes and dendritic cells, and are characterized by relatively specific ligand-receptor pairs [19,22]. It is believed that homeostatic chemokines modulate the physiological homing of naive lymphocytes and dendritic cells to secondary lymphoid tissue, and that they are involved in highly specific migration control (qualitative control).
INFLAMMATORY CHEMOKINES AND HOMEOSTATIC CHEMOKINES
- There have been great advances in chemokine research through the elucidation of the potential pathogenic roles of inflammatory chemokines in allergic diseases and autoimmune diseases, including bronchial asthma, rheumatoid arthritis, and inflammatory bowel disease. In such diseases, chemokines and their receptors promote the onset and modify the severity of each of these diseases through the selective accumulation of leukocytes in the site of inflammation. MCP-1, a prototype of the chemokine [23,24], as well as MIP-1α and MIP-1β are the CC chemokine subfamily that are isolated from monocyte and macrophages, and have been shown to be closely linked with the development of many inflammatory diseases. The signals of CC chemokine receptors CCR1, CCR2, and CCR5 are also involved in the pathogenesis of multiple sclerosis, characterized by chronic inflammation induced by monocytes, macrophages, and T cells (Table 1) [19,22]. In acute respiratory distress syndrome, on the other hand, CXC chemokines act on neutrophils, causing a large number of neutrophils to infiltrate into the lung, thus triggering a severe acute inflammatory reaction. Furthermore, in recent years, increasing evidence has shown that chemokines and their receptors play pathogenic roles on cancer, cardiovascular disease, neurodegenerative disorder, and metabolic diseases which have led many researchers to explore the role of chemokines on various other diseases associated with chronic inflammation.
CHEMOKINE SYSTEMS AND DISEASE
- It has become increasingly evident that chemokine system also involves chronic subacute inflammation that is the common underlying condition of obesity, insulin resistance, and type 2 diabetes. The interaction of MCP-1 with its receptor CCR2 is considered pivotal in obesity-induced insulin resistance. Several groups have reported that mice with targeted deletions in the genes for Mcp-1/Ccl2 and its receptor Ccr2 have decreased ATM content, decreased inflammation in fat, and protection from high-fat (HF) diet-induced insulin resistance [25,26]. Conversely, mice overexpressing MCP-1 in adipose tissues have increased numbers of ATMs along with insulin resistance [25,27]. Therefore, the MCP-1-CCR2 axis is of central importance for promoting ATM recruitment and insulin resistance in mice. More recent studies, however, have produced conflicting results and indicated greater complexity than suggested by earlier reports. Loss of MCP-1 neither attenuates obesity-associated macrophage recruitment to WAT nor improves metabolic function, suggesting that MCP-1 is not critical for obesity-induced ATM recruitment and systemic insulin resistance [28,29]. Furthermore, although Ccr2-/- mice fed a HF diet have fewer macrophages in WAT compared with WT mice [26] CCR2 deficiency does not normalize ATM content and insulin resistance to the levels in lean animals, indicating that ATM recruitment and subsequent insulin resistance are also regulated by MCP-1-CCR2 independent signals.
- This complexity and redundancy of chemokine signaling may account for these conflicting results. In fact, other chemokine systems have also been implicated in ATM infiltration in obese mice [30-32]. Indeed, serum levels of CXCL5 produced by ATM are increased in obese mice and humans, and mice lacking CXCR2, the receptor of CXCL5, show resistance to the onset of obesity-induced disorders of glucose metabolism [32]. Moreover, previous research has suggested that the concentration of keratinocyte, which is homologous to human IL-8, increases in the adipose tissue and plasma in an obese model, and that bone marrow chimera mice with bone marrow cells from CXCR2 deficient mice show decreased obesity-induced inflammation and insulin resistance compared to controls [31]. However, additional unidentified chemokine/chemokine rereceptor pathways that may play significant roles in ATM recruitment and insulin sensitivity remain to be fully identified.
PIVOTAL ROLE OF MCP-1-CCR2 IN OBESITY-INDUCED INSULIN RESISTANCE
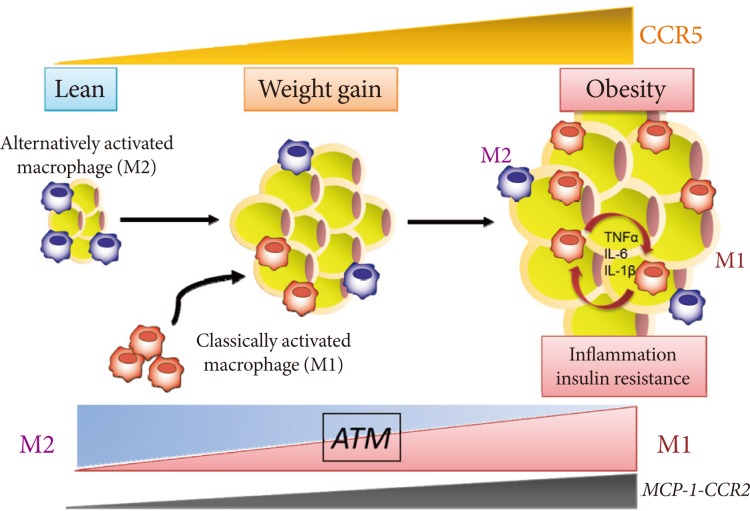
- We recently identified and characterized a critical role for CCR5, a different CC chemokine receptor, in the regulation of the adipose tissue inflammatory response to obesity and the development of insulin resistance (Fig. 1) [33]. We made several important observations. First, expression of CCR5 and its ligands is significantly increased and is equal to that of CCR2 and its ligands in the WAT of obese mice, particularly in the macrophage fraction. Second, fluorescence-activated cell sorter analysis clearly demonstrates a robust increase in CCR5+ ATMs in response to a HF diet even after normalizing for stromal vascular cell number and fat weight. Third, and most important, Ccr5-/- mice are protected from insulin resistance, hepatic steatosis, and diabetes induced by HF feeding. It is noteworthy that two distinct models, both Ccr5-/- mice and chimeric mice lacking CCR5 only in myeloid cells, are protected from HF diet-induced hyperinsulinemia and glucose intolerance through a reduction in ATM accumulation. Finally, it is interesting that an M2-dominant shift in ATM is induced in obese Ccr5-/- mice. Therefore, we conclude that deficiency of CCR5 causes an M2-dominant phenotypic shift in ATMs, which contributes to the attenuation of obesity-induced insulin resistance.
- Our study provides new information about the role of CCR5, a new chemokine system, in obesity-induced insulin resistance in an animal model [33]. It is important that the effects of CCR5 do not appear to result from global alterations in adipocyte biology. Thus, decreased ATM recruitment does not appear to be secondary to changes in adiposity because the adipocyte size of obese Ccr5-/- mice and age-matched controls is similar. Moreover, expression of adipocyte-derived factors such as leptin and adiponectin in WAT and plasma levels are similar between genotypes. Additionally, a bone marrow transplantation study revealed that lack of CCR5 expression in macrophages alone was sufficient to protect mice from the HF diet-induced insulin resistance; this was associated with a marked reduction in ATM infiltration. These data support the conclusion that CCR5+ ATMs are important in the development and maintenance of obesity-induced adipose tissue inflammation and insulin resistance in mice. Recent human studies have also shown upregulation of the expression of not only MCP-1-CCR2 but also other CC chemokines (CCL5, CCL7, CCL8, CCL11) and their receptors (CCR1, CCR3, CCR5) in the visceral fat of morbidly obese individuals in whom macrophage infiltration has been confirmed [34]. Taken together, CCR5-mediated signals in the adipose tissue may be involved, in some way, in the induction and maintenance of obesity-induced inflammation and in the development of insulin resistance in both rodents and humans.
CCR5 LINKS OBESITY TO INSULIN RESISTANCE BY REGULATING BOTH MACROPHAGE RECRUITMENT AND M1/M2 STATUS
- Do the two CC chemokine receptors, CCR2 and CCR5, play common or unique roles in obesity-induced adipose tissue inflammation and insulin resistance? Importantly, no significant compensatory increase in the expression for CCR2, or vice versa, has been found. Therefore, CCR5, independently from and/or cooperatively with CCR2, plays a role in the maintenance of ATM dysfunction and insulin resistance once obesity and its metabolic consequences have been established (Fig. 1). Moreover, similar to the case in Ccr5-/- mice, HF diet-induced increased fat mass are minimally affected by Ccr2 deficiency, and obese Ccr2-/- mice matched for adiposity with controls showed reduced ATM recruitment and improved systemic insulin sensitivity [26]. Therefore, the effects of either CCR5 or CCR2 do not appear to result from global alterations in adipocyte biology. However, HF feeding promotes accumulation of M1 macrophages in WAT of WT mice, whereas increase in M1 ATMs are markedly suppressed in Ccr5-/- mice [33]. In contrast, M2 expression within ATMs is increased in Ccr5-/- mice on a HF diet, suggesting that deficiency of CCR5 causes an M2-dominant phenotypic shift in ATMs, which contributes to the attenuation of obesity-induced insulin resistance. Interestingly, such a phenotypic switch is not observed in Ccr2-/- mice [17].
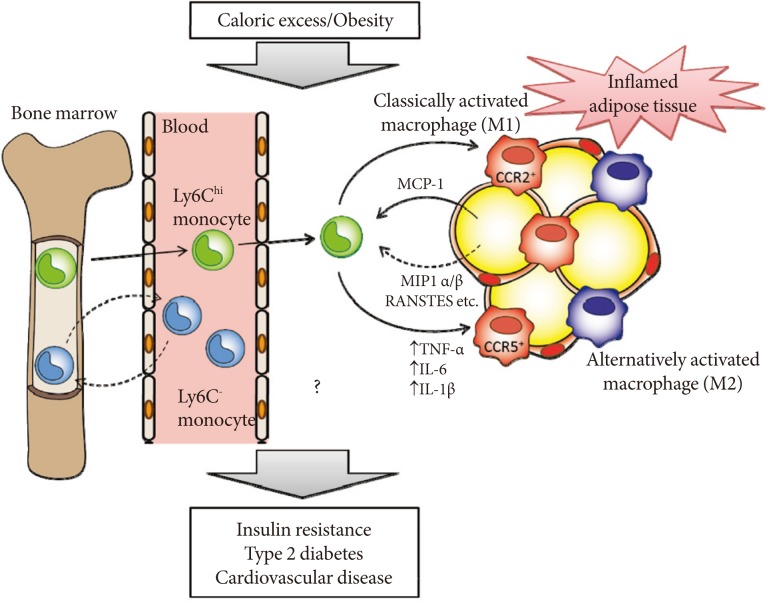
- CCR5 is preferentially expressed on Th1 cells [35]. Recent studies have demonstrated that obesity is associated with increased accumulation of not only macrophages but also T cells in adipose tissue. Wu et al. [36], showed that RANTES/CCL5 mRNA levels are highly correlated with the T cell marker CD3 in human visceral adipose tissue. However, the numbers of CD3+ T cells, CD4+ T cells and CD8+ T cells did not differ in WAT of HF diet-fed WT and Ccr5-/- mice [33], suggesting that CCR5 deficiency affects ATM recruitment more prominently. One important question concerns whether the loss of CCR5 affects the M1/M2 status in the bone marrow or peripheral blood (Fig. 2). In mice, two major distinct subsets of blood monocytes have been reported: Ly6Chi and Ly6C- monocytes. The former, called proinflammatory/classical monocytes, preferentially accumulate in atherosclerotic plaques and exhibit a strong inflammatory response to lipopolysaccharide [37]. In contrast, the latter, known as resident/remodeling/patrolling monocytes, participate in the resolution of inflammation [38]. Both Ly6Chi and Ly6C- monocytes are recruited to sites of inflammation or injury (Fig. 2) [39]. Although the relationship between the monocyte subtypes and their fate as M1/M2 macrophages remains unknown, that loss of CCR5 could cause alteration of Ly6Chi and Ly6C- monocytes subsets at the level of either bone marrow or peripheral blood, and that this contributes to the M2-dominant shift of ATM in obese Ccr5-/- mice.
- Our study provides new information regarding the role of CCR5 as a novel link among obesity, adipose tissue inflammation, and insulin resistance (Fig. 1). However, many questions have yet to be answered, including how CCR5 and its ligands are induced in response to either a HF diet or obesity; how CCR5 regulates M2 macrophages; which metabolic tissue/organ is responsible for enhanced insulin sensitivity in Ccr5-/-; and of the 50 chemokines in metabolic diseases, what distinct roles are played by CCR5? As cytokines and chemokines work in networks, the effect of individual molecules on metabolic function depends on their place in the hierarchy of the network. Several recent reports suggest that obesity increases macrophage infiltration to insulin target organs other than adipose tissue. For example, MCP-1-mediated infiltration of CCR2+ bone marrow cells was observed in the liver of the ob/ob mice, and adenovirus-induced overexpression of MCP-1 in the liver of wild-type mice treated with HF diet was found to increase lipogenesis and to promote fatty liver disease [40]. Chemokine-mediated macrophage infiltration into the liver might be therefore related with the pathology of not only insulin resistance but also steatohepatitis. In light of these new data, however, CCR5 may be a promising therapeutic target for insulin resistance, metabolic syndrome, and type 2 diabetes. Further work is required to gain a systematic understanding of how CCR5 and MCP-1-CCR2 as well as other chemokine systems, connect obesity, inflammation, and insulin resistance.
CCR5 AS A NOVEL PLAYER IN THE ADIPOSE TISSUE INFLAMMATION AND INSULIN RESISTANCE
- Vigorous research has been conducted to explore the physiological functions of chemokines in controlling inflammatory and immune response, as well as to elucidate their pathophysiological roles in the development of autoimmune diseases and allergic diseases. It has also become increasingly evident that chemokines play an important role in obesity-induced insulin resistance and its comorbidities, including type 2 diabetes and cardiovascular disease. MCP-1-CCR2 as well as other chemokine systems may be deeply involved not only in tissue- and organ-level inflammation caused by interaction between adipose tissue and macrophages, but also in the onset of localized inflammation caused by cross-talk between adipose tissue and other organs and the subsequent development of systemic insulin resistance. There are many questions, however, that have yet to be answered, including how chemokine expression is regulated in obesity, how chemokines regulate macrophage polarization, and what the different roles of over 50 chemokines are in metabolic diseases. Further research on chemokines in relation to the molecular bases of metabolic inflammation and pathogenic roles in insulin resistance is expected to contribute to the development of new drugs that could control inflammation- or immune-mediated disorders such as metabolic syndrome and type 2 diabetes.
CONCLUSIONS
-
Acknowledgements
- This work was supported by the following grants to Tsuguhito Ota: Program for Improvement of Research Environment for Young Researchers from the Special Coordination Funds for Promoting Science and Technology; Grant-in-Aid for Young Scientists (B) (22790854) from the Ministry of Education, Culture, Sports, Science and Technology of Japan; Uehara Memorial Foundation; Kanae Foundation for the Promotion of Medical Science; Japan Heart Foundation/Novartis Grant for Research Award on Molecular and Cellular Cardiology; Banyu Life Science Foundation International; Research Grant of Astellas Foundation for Research on Metabolic Disorders; Japan Research Foundation for Clinical Pharmacology and Daiichi-Sankyo Foundation of Life Science. The author thanks Dr. Mayumi Nagashimada for drawing the figure.
ACKNOWLEDGMENTS
- 1. Hotamisligil GS, Erbay E. Nutrient sensing and inflammation in metabolic diseases. Nat Rev Immunol 2008;8:923-934. ArticlePubMedPMCPDF
- 2. Hansson GK, Libby P. The immune response in atherosclerosis: a double-edged sword. Nat Rev Immunol 2006;6:508-519. ArticlePubMedPDF
- 3. Wajchenberg BL. Subcutaneous and visceral adipose tissue: their relation to the metabolic syndrome. Endocr Rev 2000;21:697-738. ArticlePubMed
- 4. Mantovani A, Allavena P, Sica A, Balkwill F. Cancer-related inflammation. Nature 2008;454:436-444. ArticlePubMedPDF
- 5. Lehr S, Hartwig S, Sell H. Adipokines: a treasure trove for the discovery of biomarkers for metabolic disorders. Proteomics Clin Appl 2012;6:91-101. ArticlePubMedPDF
- 6. Siiteri PK. Adipose tissue as a source of hormones. Am J Clin Nutr 1987;45(1 Suppl):277-282. ArticlePubMed
- 7. Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. Positional cloning of the mouse obese gene and its human homologue. Nature 1994;372:425-432. ArticlePubMedPDF
- 8. Kadowaki T, Yamauchi T, Kubota N, Hara K, Ueki K, Tobe K. Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J Clin Invest 2006;116:1784-1792. ArticlePubMedPMC
- 9. Hotamisligil GS. Inflammation and metabolic disorders. Nature 2006;444:860-867. ArticlePubMedPDF
- 10. Shoelson SE, Lee J, Goldfine AB. Inflammation and insulin resistance. J Clin Invest 2006;116:1793-1801. ArticlePubMedPMC
- 11. Hotamisligil GS, Shargill NS, Spiegelman BM. Adipose expression of tumor necrosis factor-alpha: direct role in obesity-linked insulin resistance. Science 1993;259:87-91. ArticlePubMed
- 12. Neels JG, Olefsky JM. Inflamed fat: what starts the fire? J Clin Invest 2006;116:33-35. ArticlePubMedPMC
- 13. Cancello R, Henegar C, Viguerie N, Taleb S, Poitou C, Rouault C, Coupaye M, Pelloux V, Hugol D, Bouillot JL, Bouloumie A, Barbatelli G, Cinti S, Svensson PA, Barsh GS, Zucker JD, Basdevant A, Langin D, Clement K. Reduction of macrophage infiltration and chemoattractant gene expression changes in white adipose tissue of morbidly obese subjects after surgery-induced weight loss. Diabetes 2005;54:2277-2286. ArticlePubMedPDF
- 14. Xu H, Barnes GT, Yang Q, Tan G, Yang D, Chou CJ, Sole J, Nichols A, Ross JS, Tartaglia LA, Chen H. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J Clin Invest 2003;112:1821-1830. ArticlePubMedPMC
- 15. Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante AW Jr. Obesity is associated with macrophage accumulation in adipose tissue. J Clin Invest 2003;112:1796-1808. ArticlePubMedPMC
- 16. Mantovani A, Sica A, Sozzani S, Allavena P, Vecchi A, Locati M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol 2004;25:677-686. ArticlePubMed
- 17. Lumeng CN, Bodzin JL, Saltiel AR. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J Clin Invest 2007;117:175-184. ArticlePubMedPMC
- 18. Odegaard JI, Ricardo-Gonzalez RR, Goforth MH, Morel CR, Subramanian V, Mukundan L, Eagle AR, Vats D, Brombacher F, Ferrante AW, Chawla A. Macrophage-specific PPARgamma controls alternative activation and improves insulin resistance. Nature 2007;447:1116-1120. ArticlePubMedPMCPDF
- 19. Proudfoot AE. Chemokine receptors: multifaceted therapeutic targets. Nat Rev Immunol 2002;2:106-115. ArticlePubMedPMCPDF
- 20. Baggiolini M. Chemokines and leukocyte traffic. Nature 1998;392:565-568. ArticlePubMedPDF
- 21. Gerard C, Rollins BJ. Chemokines and disease. Nat Immunol 2001;2:108-115. ArticlePubMedPDF
- 22. Zlotnik A, Yoshie O. Chemokines: a new classification system and their role in immunity. Immunity 2000;12:121-127. ArticlePubMed
- 23. Yoshimura T, Robinson EA, Tanaka S, Appella E, Kuratsu J, Leonard EJ. Purification and amino acid analysis of two human glioma-derived monocyte chemoattractants. J Exp Med 1989;169:1449-1459. ArticlePubMedPMCPDF
- 24. Matsushima K, Larsen CG, DuBois GC, Oppenheim JJ. Purification and characterization of a novel monocyte chemotactic and activating factor produced by a human myelomonocytic cell line. J Exp Med 1989;169:1485-1490. ArticlePubMedPMCPDF
- 25. Kanda H, Tateya S, Tamori Y, Kotani K, Hiasa K, Kitazawa R, Kitazawa S, Miyachi H, Maeda S, Egashira K, Kasuga M. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J Clin Invest 2006;116:1494-1505. ArticlePubMedPMC
- 26. Weisberg SP, Hunter D, Huber R, Lemieux J, Slaymaker S, Vaddi K, Charo I, Leibel RL, Ferrante AW Jr. CCR2 modulates inflammatory and metabolic effects of high-fat feeding. J Clin Invest 2006;116:115-124. ArticlePubMed
- 27. Kamei N, Tobe K, Suzuki R, Ohsugi M, Watanabe T, Kubota N, Ohtsuka-Kowatari N, Kumagai K, Sakamoto K, Kobayashi M, Yamauchi T, Ueki K, Oishi Y, Nishimura S, Manabe I, Hashimoto H, Ohnishi Y, Ogata H, Tokuyama K, Tsunoda M, Ide T, Murakami K, Nagai R, Kadowaki T. Overexpression of monocyte chemoattractant protein-1 in adipose tissues causes macrophage recruitment and insulin resistance. J Biol Chem 2006;281:26602-26614. ArticlePubMed
- 28. Inouye KE, Shi H, Howard JK, Daly CH, Lord GM, Rollins BJ, Flier JS. Absence of CC chemokine ligand 2 does not limit obesity-associated infiltration of macrophages into adipose tissue. Diabetes 2007;56:2242-2250. ArticlePubMedPDF
- 29. Kirk EA, Sagawa ZK, McDonald TO, O'Brien KD, Heinecke JW. Monocyte chemoattractant protein deficiency fails to restrain macrophage infiltration into adipose tissue [corrected]. Diabetes 2008;57:1254-1261. PubMed
- 30. Nara N, Nakayama Y, Okamoto S, Tamura H, Kiyono M, Muraoka M, Tanaka K, Taya C, Shitara H, Ishii R, Yonekawa H, Minokoshi Y, Hara T. Disruption of CXC motif chemokine ligand-14 in mice ameliorates obesity-induced insulin resistance. J Biol Chem 2007;282:30794-30803. ArticlePubMed
- 31. Neels JG, Badeanlou L, Hester KD, Samad F. Keratinocyte-derived chemokine in obesity: expression, regulation, and role in adipose macrophage infiltration and glucose homeostasis. J Biol Chem 2009;284:20692-20698. PubMedPMC
- 32. Chavey C, Lazennec G, Lagarrigue S, Clape C, Iankova I, Teyssier J, Annicotte JS, Schmidt J, Mataki C, Yamamoto H, Sanches R, Guma A, Stich V, Vitkova M, Jardin-Watelet B, Renard E, Strieter R, Tuthill A, Hotamisligil GS, Vidal-Puig A, Zorzano A, Langin D, Fajas L. CXC ligand 5 is an adipose-tissue derived factor that links obesity to insulin resistance. Cell Metab 2009;9:339-349. ArticlePubMedPMC
- 33. Kitade H, Sawamoto K, Nagashimada M, Inoue H, Yamamoto Y, Sai Y, Takamura T, Yamamoto H, Miyamoto K, Ginsberg HN, Mukaida N, Kaneko S, Ota T. CCR5 plays a critical role in obesity-induced adipose tissue inflammation and insulin resistance by regulating both macrophage recruitment and M1/M2 status. Diabetes 2012;61:1680-1690. ArticlePubMedPMCPDF
- 34. Huber J, Kiefer FW, Zeyda M, Ludvik B, Silberhumer GR, Prager G, Zlabinger GJ, Stulnig TM. CC chemokine and CC chemokine receptor profiles in visceral and subcutaneous adipose tissue are altered in human obesity. J Clin Endocrinol Metab 2008;93:3215-3221. ArticlePubMed
- 35. Bonecchi R, Bianchi G, Bordignon PP, D'Ambrosio D, Lang R, Borsatti A, Sozzani S, Allavena P, Gray PA, Mantovani A, Sinigaglia F. Differential expression of chemokine receptors and chemotactic responsiveness of type 1 T helper cells (Th1s) and Th2s. J Exp Med 1998;187:129-134. ArticlePubMedPMCPDF
- 36. Wu H, Ghosh S, Perrard XD, Feng L, Garcia GE, Perrard JL, Sweeney JF, Peterson LE, Chan L, Smith CW, Ballantyne CM. T-cell accumulation and regulated on activation, normal T cell expressed and secreted upregulation in adipose tissue in obesity. Circulation 2007;115:1029-1038. ArticlePubMed
- 37. Swirski FK, Libby P, Aikawa E, Alcaide P, Luscinskas FW, Weissleder R, Pittet MJ. Ly-6Chi monocytes dominate hypercholesterolemia-associated monocytosis and give rise to macrophages in atheromata. J Clin Invest 2007;117:195-205. ArticlePubMedPMC
- 38. Nahrendorf M, Swirski FK, Aikawa E, Stangenberg L, Wurdinger T, Figueiredo JL, Libby P, Weissleder R, Pittet MJ. The healing myocardium sequentially mobilizes two monocyte subsets with divergent and complementary functions. J Exp Med 2007;204:3037-3047. ArticlePubMedPMCPDF
- 39. Auffray C, Fogg D, Garfa M, Elain G, Join-Lambert O, Kayal S, Sarnacki S, Cumano A, Lauvau G, Geissmann F. Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science 2007;317:666-670. ArticlePubMed
- 40. Obstfeld AE, Sugaru E, Thearle M, Francisco AM, Gayet C, Ginsberg HN, Ables EV, Ferrante AW Jr. C-C chemokine receptor 2 (CCR2) regulates the hepatic recruitment of myeloid cells that promote obesity-induced hepatic steatosis. Diabetes 2010;59:916-925. ArticlePubMedPMCPDF
REFERENCES
Fig. 1C-C motif chemokine receptor 5 (CCR5) promotes obesity-induced inflammation and insulin resistance. Kitade et al. [33] recently identified and characterized a role for CCR5, a CC chemokine receptor, and made several important observations. First, expression of CCR5 and its ligands is significantly increased and is equal to that of CCR2 and its ligands in white adipose tissue (WAT) of obese mice. Second, CCR5+ adipose tissue macrophages (ATMs) accumulate in WAT of obese mice. Third, Ccr5-/- mice are protected from insulin resistance and diabetes through both reduction in ATM accumulation and induction of an alternatively activated, M2 dominant shift in ATM. Taken together, these data indicate that CCR5 provides a novel link between obesity, adipose tissue inflammation, and insulin resistance. TNF, tumor necrosis factor; IL, interleukin; MCP, monocyte chemoattractant protein.
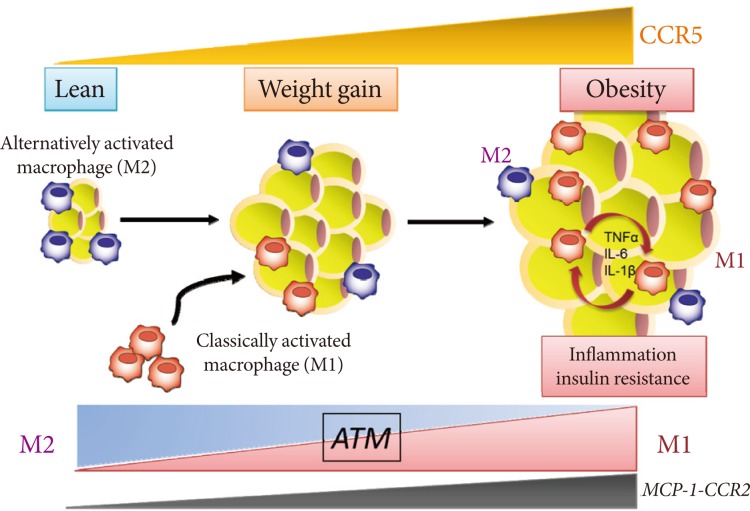
Fig. 2Hypothetical role of C-C motif chemokine receptor 5 (CCR5) and monocyte chemoattractant protein (MCP)-1-CCR2 in the development of adipose tissue inflammation and insulin resistance in obesity. Obesity causes both the recruitment and proiflammatory activation of adipose tissue macrophages (ATMs). Adipocytes or preadipocytes begin to secrete MCP-1 as well as other chemokines, such as MIP1α, MIP1β, and RANSTES (ligands for CCR5) in obesity. Thereafter, CCR2+ and/or CCR5+ macrophages accumulate and presumably maintain the inflammation as M1 or classically activated macrophages in obese adipose tissue. Ly6Chi monocytes exit the bone marrow in a CCR2-dependent manner and are recruited to inflamed tissues. CCR5 may also regulate recruitment of Ly6Chi and Ly6C- monocytes and their fate as M1/M2 ATMs. Once these ATMs are present and active, cytokines (tumor necrosis factor [TNF]-α, interleukin [IL]-6, and IL-1β) are produced. Therefore, CCR5, independently from and/or cooperatively with CCR2, could play an important role in the induction and maintenance of obesity-induced inflammation and insulin resistance.
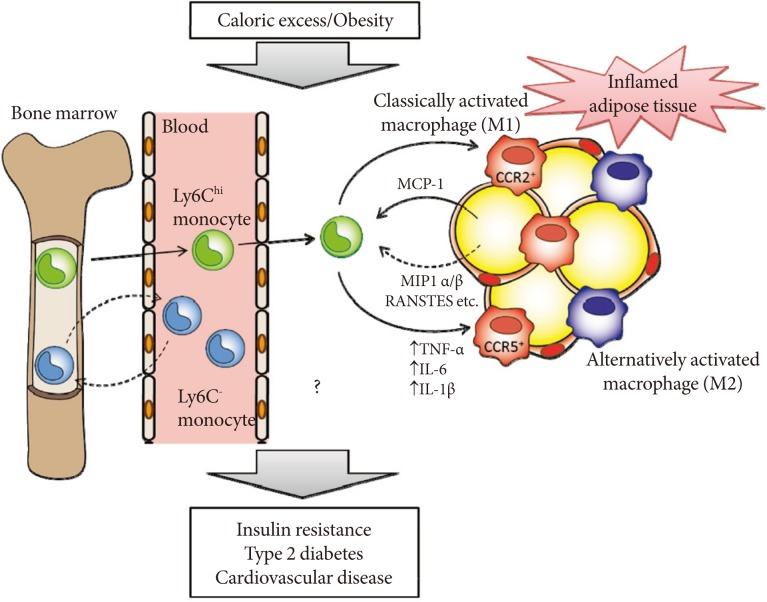
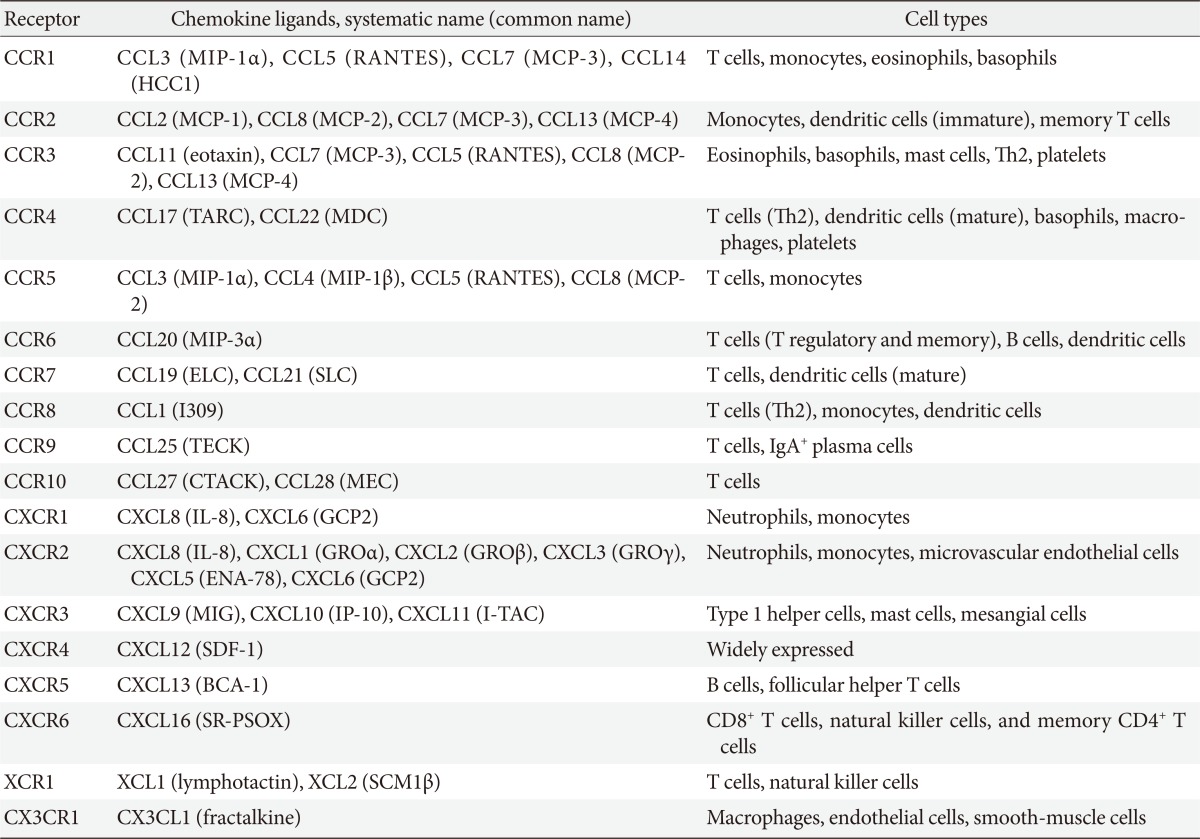
Figure & Data
References
Citations
Citations to this article as recorded by 
- Apolipoprotein A-IV reduced metabolic inflammation in white adipose tissue by inhibiting IKK and JNK signaling in adipocytes
Xiao-Huan Liu, Yupeng Zhang, Liao Chang, Yang Wei, Na Huang, Jin-Ting Zhou, Cheng Cheng, Jianbo Zhang, Jing Xu, Zongfang Li, Xiaoming Li
Molecular and Cellular Endocrinology.2023; 559: 111813. CrossRef - Neutralizing MIP3αReduces Renal Immune Cell Infiltration and Progressive Renal Injury in Young Obese Dahl Salt-Sensitive Rats
Ubong S. Ekperikpe, Bibek Poudel, Corbin A. Shields, Sautan Mandal, Denise C. Cornelius, Jan M. Williams
Journal of Pharmacology and Experimental Therapeutics.2023; 384(3): 445. CrossRef - Metformin reduces insulin resistance and attenuates progressive renal injury in prepubertal obese Dahl salt-sensitive rats
Ubong S. Ekperikpe, Sautan Mandal, Stephen J. Holt, Jacori K. Daniels, Tyler D. Johnson, Jonita S. Cooper, Sarah M. Safir, Denise C. Cornelius, Jan M. Williams
American Journal of Physiology-Renal Physiology.2023; 325(3): F363. CrossRef - Hepatic T-cell senescence and exhaustion are implicated in the progression of fatty liver disease in patients with type 2 diabetes and mouse model with nonalcoholic steatohepatitis
Byeong Chang Sim, Yea Eun Kang, Sun Kyoung You, Seong Eun Lee, Ha Thi Nga, Ho Yeop Lee, Thi Linh Nguyen, Ji Sun Moon, Jingwen Tian, Hyo Ju Jang, Jeong Eun Lee, Hyon-Seung Yi
Cell Death & Disease.2023;[Epub] CrossRef - Inflammatory Status of Monocytes in Type 2 Diabetes Mellitus
T. V. Kirichenko, L. A. Bochkareva, L. V. Nedosugova, Yu. V. Markina, I. A. Kuzina, N. A. Petunina, T. V. Tolstik, A. I. Bogatyreva, V. A. Antonov, A. M. Markin
Journal Biomed.2023; 19(4): 25. CrossRef - Non-alcoholic fatty liver disease: a multi-system disease influenced by ageing and sex, and affected by adipose tissue and intestinal function
Josh Bilson, Jaswinder K. Sethi, Christopher D. Byrne
Proceedings of the Nutrition Society.2022; 81(2): 146. CrossRef - Puerarin Attenuates Obesity-Induced Inflammation and Dyslipidemia by Regulating Macrophages and TNF-Alpha in Obese Mice
Ji-Won Noh, Hee-Kwon Yang, Min-Soo Jun, Byung-Cheol Lee
Biomedicines.2022; 10(1): 175. CrossRef - Beneficial effects of fermented barley extracts on inflammatory status and gut microbiota in high-fat diet-induced obese rats
Xiaozhong Huang, Qingguo Cao, Cen Chen, Zhenglin Xie, Jinshan Fan
Journal of Applied Microbiology.2022; 133(6): 3708. CrossRef - Periodontitis induced by Porphyromonas gingivalis drives impaired glucose metabolism in mice
Ni Kang, Yong Zhang, Fei Xue, Jinyu Duan, Fan Chen, Yu Cai, Qingxian Luan
Frontiers in Cellular and Infection Microbiology.2022;[Epub] CrossRef - Granulocytic MDSC with Deficient CCR5 Alleviates Lipogenesis and Inflammation in Nonalcoholic Fatty Liver Disease
Tzu-Chieh Liao, Jiung-Pang Huang, Yu-Ting Tsai, Wei-Ching Shih, Chi-Chang Juan, Po-Shiuan Hsieh, Li-Man Hung, Chao-Lan Yu
International Journal of Molecular Sciences.2022; 23(21): 13048. CrossRef - Metabolic and proteomic signatures of type 2 diabetes subtypes in an Arab population
Shaza B. Zaghlool, Anna Halama, Nisha Stephan, Valborg Gudmundsdottir, Vilmundur Gudnason, Lori L. Jennings, Manonanthini Thangam, Emma Ahlqvist, Rayaz A. Malik, Omar M. E. Albagha, Abdul Badi Abou‑Samra, Karsten Suhre
Nature Communications.2022;[Epub] CrossRef - CX3CL1-CX3CR1 Signaling Deficiency Exacerbates Obesity-induced Inflammation and Insulin Resistance in Male Mice
Mayumi Nagashimada, Kazuki Sawamoto, Yinhua Ni, Hironori Kitade, Naoto Nagata, Liang Xu, Masuko Kobori, Naofumi Mukaida, Tatsuya Yamashita, Shuichi Kaneko, Tsuguhito Ota
Endocrinology.2021;[Epub] CrossRef - Di(2-ethylhexyl)phthalate exposure exacerbates metabolic disorders in diet-induced obese mice
Jhih-Wei Hsu, Chung-Yi Nien, Hsin-Wei Chen, Feng-Yuan Tsai, Szu-Ching Yeh, Yung-Hsi Kao, Tsui-Chun Tsou
Food and Chemical Toxicology.2021; 156: 112439. CrossRef - The Chemokine Systems at the Crossroads of Inflammation and Energy Metabolism in the Development of Obesity
Pei-Chi Chan, Po-Shiuan Hsieh
International Journal of Molecular Sciences.2021; 22(24): 13528. CrossRef - β-Neoendorphin Enhances Wound Healing by Promoting Cell Migration in Keratinocyte
Dong Joo Yang, Sang Hyun Moh, Yun-Hee Choi, Ki Woo Kim
Molecules.2020; 25(20): 4640. CrossRef - Transcriptome Analysis of Testis from HFD-Induced Obese Rats (Rattus norvigicus) Indicated Predisposition for Male Infertility
Ahmed M. El-Shehawi, Samir El-Shazly, Mohamed Ahmed, Mohamed Alkafafy, Samy Sayed, Samy Farouk, Saqer S. Alotaibi, Mona M. Elseehy
International Journal of Molecular Sciences.2020; 21(18): 6493. CrossRef - Anti-Obesity Effects of Petasites japonicus (Meowi) Ethanol Extract on RAW 264.7 Macrophages and 3T3-L1 Adipocytes and Its Characterization of Polyphenolic Compounds
Eun Mi Ahn, Gelila Asamenew, Heon Woong Kim, Sang Hoon Lee, Seon-Mi Yoo, Soo-Muk Cho, Youn-Soo Cha, Min-Sook Kang
Nutrients.2020; 12(5): 1261. CrossRef - Polymorphisms in miRNA binding sites involved in metabolic diseases in mice and humans
Pascal Gottmann, Meriem Ouni, Lisa Zellner, Markus Jähnert, Kilian Rittig, Dirk Walther, Annette Schürmann
Scientific Reports.2020;[Epub] CrossRef - Asthma and metabolic syndrome: a comprehensive systematic review and meta-analysis of observational studies
Nahid Karamzad, Neda Izadi, Sarvin Sanaie, Elham Ahmadian, Aziz Eftekhari, Mark J.M. Sullman, Saeid Safiri
Journal of Cardiovascular and Thoracic Research.2020; 12(2): 120. CrossRef - Elevated adipose tissue associated IL-2 expression in obesity correlates with metabolic inflammation and insulin resistance
Shihab Kochumon, Ashraf Al Madhoun, Fatema Al-Rashed, Reeby Thomas, Sardar Sindhu, Ebaa Al-Ozairi, Fahd Al-Mulla, Rasheed Ahmad
Scientific Reports.2020;[Epub] CrossRef - Obesity-related hypoxia via miR-128 decreases insulin-receptor expression in human and mouse adipose tissue promoting systemic insulin resistance
Biagio Arcidiacono, Eusebio Chiefari, Anna Foryst-Ludwig, Giuseppe Currò, Giuseppe Navarra, Francesco S. Brunetti, Maria Mirabelli, Domenica M. Corigliano, Ulrich Kintscher, Domenico Britti, Vincenzo Mollace, Daniela P. Foti, Ira D. Goldfine, Antonio Brun
eBioMedicine.2020; 59: 102912. CrossRef - Chemokines and their association with body mass index among healthy Saudis
Steve Harakeh, Gauthaman Kalamegam, Peter N. Pushparaj, Ahmed Al-Hejin, Sulaiman M. Alfadul, Turki Al Amri, Salah Barnawi, Hadeel Al Sadoun, Ahmed A. Mirza, Esam Azhar
Saudi Journal of Biological Sciences.2020; 27(1): 6. CrossRef - New Insights Into Implications of CTRP3 in Obesity, Metabolic Dysfunction, and Cardiovascular Diseases: Potential of Therapeutic Interventions
Bei Guo, Tongtian Zhuang, Feng Xu, Xiao Lin, Fuxingzi Li, Su-Kang Shan, Feng Wu, Jia-Yu Zhong, Yi Wang, Ming-Hui Zheng, Qiu-Shuang Xu, Ullah Muhammad Hasnain Ehsan, Ling-Qing Yuan
Frontiers in Physiology.2020;[Epub] CrossRef - Contribution of Oxidative Stress and Impaired Biogenesis of Pancreatic β-Cells to Type 2 Diabetes
Petr Ježek, Martin Jabůrek, Lydie Plecitá-Hlavatá
Antioxidants & Redox Signaling.2019; 31(10): 722. CrossRef - Interleukin-22 Induces the Infiltration of Visceral Fat Tissue by a Discrete Subset of Duffy Antigen Receptor for Chemokine-Positive M2-Like Macrophages in Response to a High Fat Diet
Eun-Young Kim, Hye Mi Noh, Bongkun Choi, Ji-Eun Park, Ji-Eun Kim, Youngsaeng Jang, Hyung Keun Lee, Eun-Ju Chang
Cells.2019; 8(12): 1587. CrossRef - The chemokine system and its role in obesity
Wenhua Xue, Zhirui Fan, Lifeng Li, Jingli Lu, Yunkai Zhai, Jie Zhao
Journal of Cellular Physiology.2019; 234(4): 3336. CrossRef - Inonotus sanghuang Polyphenols Attenuate Inflammatory Response Via Modulating the Crosstalk Between Macrophages and Adipocytes
Mengdi Zhang, Yu Xie, Xing Su, Kun Liu, Yijie Zhang, Wuyan Pang, Junpeng Wang
Frontiers in Immunology.2019;[Epub] CrossRef - Therapeutic Effects of Adipose Stem Cells from Diabetic Mice for the Treatment of Type 2 Diabetes
Mengmeng Wang, Lili Song, Charlie Strange, Xiao Dong, Hongjun Wang
Molecular Therapy.2018; 26(8): 1921. CrossRef - Inflammatory Cytokine Profiles in Visceral and Subcutaneous Adipose Tissues of Obese Patients Undergoing Bariatric Surgery Reveal Lack of Correlation With Obesity or Diabetes
Volatiana Rakotoarivelo, Gregory Lacraz, Marian Mayhue, Christine Brown, Diane Rottembourg, Julie Fradette, Subburaj Ilangumaran, Alfredo Menendez, Marie-France Langlois, Sheela Ramanathan
eBioMedicine.2018; 30: 237. CrossRef - Resveratrol reduces the inflammatory response in adipose tissue and improves adipose insulin signaling in high-fat diet-fed mice
Shibin Ding, Jinjin Jiang, Zhe Wang, Guofu Zhang, Jianli Yin, Xiaoya Wang, Sui Wang, Zengli Yu
PeerJ.2018; 6: e5173. CrossRef - Dual CCR2/5 Antagonist Attenuates Obesity‐Induced Insulin Resistance by Regulating Macrophage Recruitment and M1/M2 Status
Ji Hye Huh, Hong Min Kim, Eun Soo Lee, Mi Hye Kwon, Bo Ra Lee, Hyun‐Jeong Ko, Choon Hee Chung
Obesity.2018; 26(2): 378. CrossRef - Rosiglitazone Elicits an Adiponectin-Mediated Insulin-Sensitizing Action at the Adipose Tissue-Liver Axis in Otsuka Long-Evans Tokushima Fatty Rats
Jia Li, Yao-Ming Xue, Bo Zhu, Yong-Hua Pan, Yan Zhang, Chunxia Wang, Yuhao Li
Journal of Diabetes Research.2018; 2018: 1. CrossRef - Myeloid-specific deletion of thrombospondin 1 protects against inflammation and insulin resistance in long-term diet-induced obese male mice
Hasiyet Memetimin, Dong Li, Kaiyuan Tan, Changcheng Zhou, Ying Liang, Yadi Wu, Shuxia Wang
American Journal of Physiology-Endocrinology and Metabolism.2018; 315(6): E1194. CrossRef - From chronic overnutrition to metaflammation and insulin resistance: adipose tissue and liver contributions
Tiziana Caputo, Federica Gilardi, Béatrice Desvergne
FEBS Letters.2017; 591(19): 3061. CrossRef - Effects of Karela (Bitter Melon; Momordica charantia) on genes of lipids and carbohydrates metabolism in experimental hypercholesterolemia: biochemical, molecular and histopathological study
Dalia Yossri Saad, Mohamed Mohamed Soliman, Ahmed A. Baiomy, Magdy Hassan Yassin, Hanan Basiouni El-Sawy
BMC Complementary and Alternative Medicine.2017;[Epub] CrossRef - Polygala tenuifolia extract inhibits lipid accumulation in 3T3-L1 adipocytes and high-fat diet–induced obese mouse model and affects hepatic transcriptome and gut microbiota profiles
Chun-Chung Wang, Jui-Hung Yen, Yi-Cheng Cheng, Chia-Yu Lin, Cheng-Ta Hsieh, Rung-Jiun Gau, Shu-Jiau Chiou, Hwan-You Chang
Food & Nutrition Research.2017; 61(1): 1379861. CrossRef - Inflammation and the Metabolic Syndrome: The Tissue-Specific Functions of NF-κB
Leen Catrysse, Geert van Loo
Trends in Cell Biology.2017; 27(6): 417. CrossRef - RETRACTED ARTICLE: TREM-1 associated macrophage polarization plays a significant role in inducing insulin resistance in obese population
Saravanan Subramanian, Pradeep K. Pallati, Poonam Sharma, Devendra K. Agrawal, Kalyana C. Nandipati
Journal of Translational Medicine.2017;[Epub] CrossRef - The Role of Heparin Cofactor Ⅱ in the Regulation of Insulin Sensitivity and Maintenance of Glucose Homeostasis in Humans and Mice
Kiyoe Kurahashi, Seika Inoue, Sumiko Yoshida, Yasumasa Ikeda, Kana Morimoto, Ryoko Uemoto, Kazue Ishikawa, Takeshi Kondo, Tomoyuki Yuasa, Itsuro Endo, Masato Miyake, Seiichi Oyadomari, Toshio Matsumoto, Masahiro Abe, Hiroshi Sakaue, Ken-ichi Aihara
Journal of Atherosclerosis and Thrombosis.2017; 24(12): 1215. CrossRef - Pure total flavonoids from citrus improve non-alcoholic fatty liver disease by regulating TLR/CCL signaling pathway: A preliminary high-throughput ‘omics’ study
Liyan Wu, Maoxiang Yan, Jianping Jiang, Beihui He, Wei Hong, Zhiyun Chen
Biomedicine & Pharmacotherapy.2017; 93: 316. CrossRef - The association between metabolic health, obesity phenotype and the risk of breast cancer
Yong‐Moon Mark Park, Alexandra J. White, Hazel B. Nichols, Katie M. O'Brien, Clarice R. Weinberg, Dale P. Sandler
International Journal of Cancer.2017; 140(12): 2657. CrossRef - Immune regulation of glucose and lipid metabolism
Tsuguhito Ota
Diabetology International.2017; 8(3): 257. CrossRef - Distinguishing features of body mass index and psoriasis in men and women in Japan: A hospital‐based case–control study
Reiko Naito, Shinichi Imafuku
The Journal of Dermatology.2016; 43(12): 1406. CrossRef - Change in Serum Bilirubin Level as a Predictor of Incident Metabolic Syndrome
You-Bin Lee, Seung-Eun Lee, Ji Eun Jun, Jae Hwan Jee, Ji Cheol Bae, Sang-Man Jin, Jae Hyeon Kim, Sheng-Nan Lu
PLOS ONE.2016; 11(12): e0168253. CrossRef - Mechanisms of inflammatory responses and development of insulin resistance: how are they interlinked?
Kanwal Rehman, Muhammad Sajid Hamid Akash
Journal of Biomedical Science.2016;[Epub] CrossRef - Chemokine genetic polymorphism in human health and disease
Tabish Qidwai
Immunology Letters.2016; 176: 128. CrossRef - The P72R Polymorphism of p53 Predisposes to Obesity and Metabolic Dysfunction
Che-Pei Kung, Julia I-Ju Leu, Subhasree Basu, Sakina Khaku, Frederick Anokye-Danso, Qin Liu, Donna L. George, Rexford S. Ahima, Maureen E. Murphy
Cell Reports.2016; 14(10): 2413. CrossRef - E3 ubiquitin ligases as novel targets for inflammatory diseases
Santosh Kumar Goru, Anuradha Pandey, Anil Bhanudas Gaikwad
Pharmacological Research.2016; 106: 1. CrossRef - Immune and Inflammatory Signaling Pathways in Exercise and Obesity
Karsten Krüger, Frank C. Mooren, Klaus Eder, Robert Ringseis
American Journal of Lifestyle Medicine.2016; 10(4): 268. CrossRef - CCL2 level is elevated with metabolic syndrome and CXCL10 level is correlated with visceral fat area in obese children
Masahiro Ishii, Shunsuke Araki, Motohide Goto, Yukiyo Yamamoto, Koichi Kusuhara
Endocrine Journal.2016; 63(9): 795. CrossRef - CCL2 gene polymorphism is associated with post-transplant diabetes mellitus
Ewa Dabrowska-Zamojcin, Maciej Romanowski, Violetta Dziedziejko, Agnieszka Maciejewska-Karlowska, Marek Sawczuk, Krzysztof Safranow, Leszek Domanski, Andrzej Pawlik
International Immunopharmacology.2016; 32: 62. CrossRef - Blocking CXCR7-mediated adipose tissue macrophages chemotaxis attenuates insulin resistance and inflammation in obesity
Hongxia Peng, Hu Zhang, Honglei Zhu
Biochemical and Biophysical Research Communications.2016; 479(4): 649. CrossRef - A brief glimpse at CTRP3 and CTRP9 in lipid metabolism and cardiovascular protection
Yang Yang, Yue Li, Zhiqiang Ma, Shuai Jiang, Chongxi Fan, Wei Hu, Dongjin Wang, Shouyin Di, Yang Sun, Wei Yi
Progress in Lipid Research.2016; 64: 170. CrossRef - Obesity-associated cancer: an immunological perspective
Melissa J. Conroy, Margaret R. Dunne, Claire L. Donohoe, John V. Reynolds
Proceedings of the Nutrition Society.2016; 75(2): 125. CrossRef - The Dietary Isoflavone Daidzein Reduces Expression of Pro-Inflammatory Genes through PPARα/γ and JNK Pathways in Adipocyte and Macrophage Co-Cultures
Yuri Sakamoto, Junko Kanatsu, Mariko Toh, Ayano Naka, Kazuo Kondo, Kaoruko Iida, Makoto Makishima
PLOS ONE.2016; 11(2): e0149676. CrossRef - Response of gut microbiota and inflammatory status to bitter melon (Momordica charantia L.) in high fat diet induced obese rats
Juan Bai, Ying Zhu, Ying Dong
Journal of Ethnopharmacology.2016; 194: 717. CrossRef - SIRT1 and insulin resistance
Yue Cao, Xinli Jiang, Huijie Ma, Yuling Wang, Peng Xue, Yan Liu
Journal of Diabetes and its Complications.2016; 30(1): 178. CrossRef - CCL2 Serum Levels and Adiposity Are Associated with the Polymorphic Phenotypes -2518A on CCL2 and 64ILE on CCR2 in a Mexican Population with Insulin Resistance
Milton-Omar Guzmán-Ornelas, Marcelo Heron Petri, Mónica Vázquez-Del Mercado, Efraín Chavarría-Ávila, Fernanda-Isadora Corona-Meraz, Sandra-Luz Ruíz-Quezada, Perla-Monserrat Madrigal-Ruíz, Jorge Castro-Albarrán, Flavio Sandoval-García, Rosa-Elena Navarro-H
Journal of Diabetes Research.2016; 2016: 1. CrossRef - Silencing CCR2 in Macrophages Alleviates Adipose Tissue Inflammation and the Associated Metabolic Syndrome in Dietary Obese Mice
Jongkil Kim, Kunho Chung, Changseon Choi, Jagadish Beloor, Irfan Ullah, Nahyeon Kim, Kuen Yong Lee, Sang-Kyung Lee, Priti Kumar
Molecular Therapy - Nucleic Acids.2016; 5: e280. CrossRef - Les adipokines : état des lieux et nouveautés
J.-P. Bastard, C. Bastard, S. Fellahi, C. Vatier, J. Capeau, B. Fève
Obésité.2016; 11(3): 181. CrossRef - TM-25659-Induced Activation of FGF21 Level Decreases Insulin Resistance and Inflammation in Skeletal Muscle via GCN2 Pathways
Jong Gab Jung, Sang-A Yi, Sung-E Choi, Yup Kang, Tae Ho Kim, Ja Young Jeon, Myung Ae Bae, Jin Hee Ahn, Hana Jeong, Eun Sook Hwang, Kwan-Woo Lee
Molecules and Cells.2015; 38(12): 1037. CrossRef - Aromatase Controls Sjögren Syndrome–Like Lesions through Monocyte Chemotactic Protein-1 in Target Organ and Adipose Tissue–Associated Macrophages
Akihiko Iwasa, Rieko Arakaki, Naoko Honma, Aya Ushio, Akiko Yamada, Tomoyuki Kondo, Emi Kurosawa, Satoko Kujiraoka, Takaaki Tsunematsu, Yasusei Kudo, Eiji Tanaka, Noriko Yoshimura, Nobuhiro Harada, Yoshio Hayashi, Naozumi Ishimaru
The American Journal of Pathology.2015; 185(1): 151. CrossRef - Macrophage Migration Inhibitory Factor Promoter Polymorphisms (−794 CATT5–8and −173 G>C): Relationship with mRNA Expression and Soluble MIF Levels in Young Obese Subjects
Inés Matia-García, Lorenzo Salgado-Goytia, José F. Muñoz-Valle, Samuel García-Arellano, Jorge Hernández-Bello, Aralia B. Salgado-Bernabé, Isela Parra-Rojas
Disease Markers.2015; 2015: 1. CrossRef - Niche-Dependent Regulations of Metabolic Balance in High-Fat Diet–Induced Diabetic Mice by Mesenchymal Stromal Cells
Andrea Tung-Qian Ji, Yun-Chuang Chang, Yun-Ju Fu, Oscar K. Lee, Jennifer H. Ho
Diabetes.2015; 64(3): 926. CrossRef - Simvastatin Does Not Reduce Chemokine Production in Obesity Without Comorbidities
Karla Silva Fernandes, Samantha Ribeiro Béla, Vanessa L. Andrade, Tatiane Figueiredo de Moraes, Olindo de Assis Martins-Filho, Valéria Cristina Sandrim
Inflammation.2015; 38(3): 1297. CrossRef - Asthma and metabolic syndrome: Current knowledge and future perspectives
Laura Serafino-Agrusa
World Journal of Clinical Cases.2015; 3(3): 285. CrossRef - Analysis of the correlation between serum resistin and the variability of erythropoietin responsiveness in patients with chronic kidney disease
HONGHAO ZHANG, XIUJIANG LI, YANHONG KAN, FAN YANG, YUE HOU, YUJUN DU
Experimental and Therapeutic Medicine.2015; 10(5): 1925. CrossRef - Preventive effects of bitter melon (Momordica charantia) against insulin resistance and diabetes are associated with the inhibition of NF-κB and JNK pathways in high-fat-fed OLETF rats
Soo Jin Yang, Jung Mook Choi, Se Eun Park, Eun Jung Rhee, Won Young Lee, Ki Won Oh, Sung Woo Park, Cheol-Young Park
The Journal of Nutritional Biochemistry.2015; 26(3): 234. CrossRef - Association of serum angiopoietin-like protein 2 with carotid intima-media thickness in subjects with type 2 diabetes
Chang Hee Jung, Woo Je Lee, Min Jung Lee, Yu Mi Kang, Jung Eun Jang, Jaechan Leem, Yoo La Lee, So Mi Seol, Hae Kyeong Yoon, Joong-Yeol Park
Cardiovascular Diabetology.2015;[Epub] CrossRef - Effects of intermittent exercise on biomarkers of cardiovascular risk in night shift workers
Seung-Taek Lim, Seok-Ki Min, Yoo-Chan Kwon, Sang-Kab Park, Hyuntae Park
Atherosclerosis.2015; 242(1): 186. CrossRef - Milk-derived peptide Val-Pro-Pro (VPP) inhibits obesity-induced adipose inflammation via an angiotensin-converting enzyme (ACE) dependent cascade
Yoko Sawada, Yuri Sakamoto, Mariko Toh, Nozomi Ohara, Yuiko Hatanaka, Ayano Naka, Yoshimi Kishimoto, Kazuo Kondo, Kaoruko Iida
Molecular Nutrition & Food Research.2015; 59(12): 2502. CrossRef - Ubiquitin ligase Cbl-b and obesity-induced insulin resistance [Review]
Tomoki Abe, Katsuya Hirasaka, Shohei Kohno, Arisa Ochi, Naoko Yamagishi, Ayako Ohno, Shigetada Teshima-Kondo, Takeshi Nikawa
Endocrine Journal.2014; 61(6): 529. CrossRef - Deficiency in adipocyte chemokine receptor CXCR4 exacerbates obesity and compromises thermoregulatory responses of brown adipose tissue in a mouse model of diet‐induced obesity
Longbiao Yao, Janet Heuser‐Baker, Oana Herlea‐Pana, Nan Zhang, Luke I. Szweda, Timothy M. Griffin, Jana Barlic‐Dicen
The FASEB Journal.2014; 28(10): 4534. CrossRef - Unprotected daily sun exposure is differently associated with central adiposity and beta-cell dysfunction by gender: The Korean national health and nutrition examination survey (KNHANES) V
Jung Hun Ohn, In Ho Kwon, Juri Park, Ohk Hyun Ryu, Seong Jin Lee, Doo-Man Kim, Sung-Hee Ihm, Moon-Gi Choi, Hyung Joon Yoo, Eun-Gyoung Hong
Environmental Research.2014; 133: 253. CrossRef - Anti-adipogenic effect of mulberry leaf ethanol extract in 3T3-L1 adipocytes
Soo Jin Yang, Na-Young Park, Yunsook Lim
Nutrition Research and Practice.2014; 8(6): 613. CrossRef - What We Talk About When We Talk About Fat
Evan D. Rosen, Bruce M. Spiegelman
Cell.2014; 156(1-2): 20. CrossRef - Effect of diabetes on the relation between troponin T and inflammatory markers in patients on hemodialysis
Emília Mácsai, Ilona Németh, Attila Benke, Gyula Dávid
Orvosi Hetilap.2014; 155(16): 627. CrossRef - Active ingredients from natural botanicals in the treatment of obesity
W.‐L. Zhang, L. Zhu, J.‐G. Jiang
Obesity Reviews.2014; 15(12): 957. CrossRef - Hypertrophic Obesity and Subcutaneous Adipose Tissue Dysfunction
Anna Meiliana, Andi Wijaya
The Indonesian Biomedical Journal.2014; 6(2): 79. CrossRef - Obesity and Cytokines in Childhood-Onset Systemic Lupus Erythematosus
Nailú Angélica Sinicato, Mariana Postal, Fernando Augusto Peres, Karina de Oliveira Peliçari, Roberto Marini, Allan de Oliveira dos Santos, Celso Dario Ramos, Simone Appenzeller
Journal of Immunology Research.2014; 2014: 1. CrossRef - Assessment of the number and function of macrophages in the placenta of gestational diabetes mellitus patients
Jun Yu, Yong Zhou, Juan Gui, Ai-zhen Li, Xiao-ling Su, Ling Feng
Journal of Huazhong University of Science and Technology [Medical Sciences].2013; 33(5): 725. CrossRef
 PubReader
PubReader Cite
Cite